

前言
Janus 激酶(JAK)-信号转导和转录激活因子 (STAT) 通路的发现源于对细胞如何响应干扰素(IFN)的研究,提供了一种细胞信号转导的范例。
美国国立卫生研究院 (NIH)的John J. O’Shea及其团队是解析JAKs和STATs家族通路在免疫调节中作用的领先者,他们综述的JAK-STAT通路研究文章于2022年10月发表在CELL期刊上,概述了JAK-STAT通路的主要里程碑式事件,并讨论了目前的重大挑战和未来机遇。

01
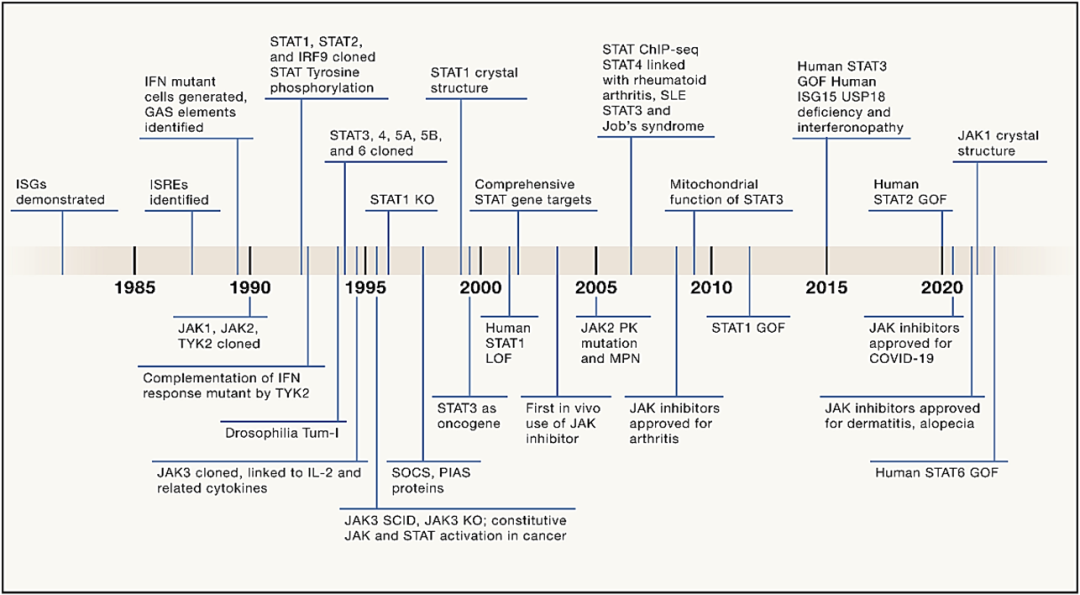
图1 JAK-STAT 时间线(自 1984 年以来JAK-STAT通路的各种里程碑)
02
信号转导的新范例
▲从IFN处理诱导的mRNA中产生cDNA文库的能力允许发现几个IFN诱导的基因,从而确定了控制IFN诱导基因表达的启动子和调控元件。
▲Stark和Kerr实验室使用IFN诱导基因的启动子,经过诱变,分离出抗性细胞克隆,通过遗传互补,产生TYK2作为诱变剂灭活的必需基因。
▲使用相对较新的简并聚合酶链式反应方法生成蛋白激酶文库,鉴定了新的 TYK 家族,包括TYK2、JAK1 和 JAK2。JAK这个名字起源于两面罗马神Janus,因为该激酶家族具有特征性的双面结构。
▲Darnell实验室纯化出STAT(STAT1和STAT2,干扰素调节因子9(IRF9),其命名表示它们的双重性功能,并强调它们的功能执行速度快速。
03
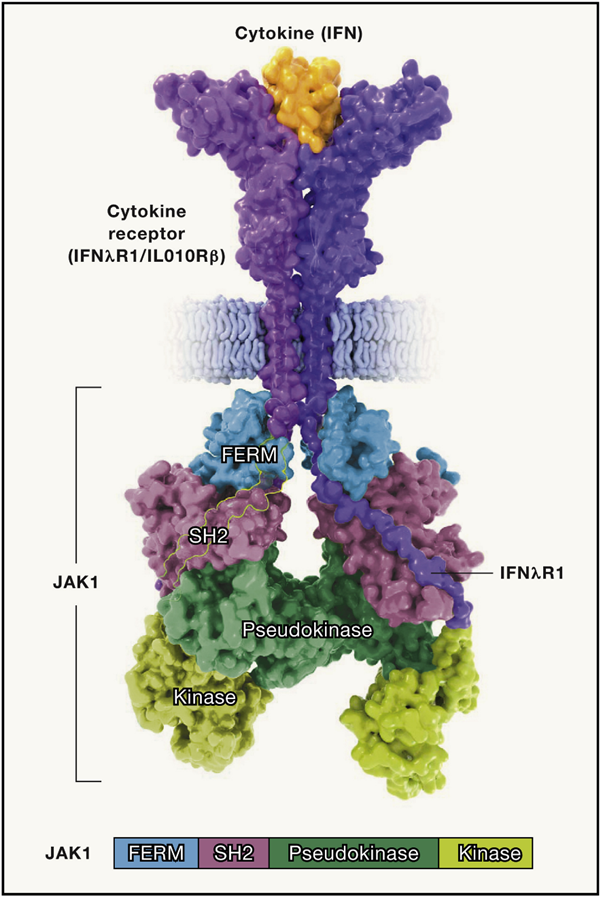
图2 Janus激酶的结构
STAT3被鉴定为响应IL-6的DNA结合“急性期反应因子”。表皮生长因子和其他生长因子也被认为是STAT3激活剂。STAT4主要在淋巴组织中表达并被IL-12、IL-23和I型干扰素激活。由催乳素激活的“乳腺特异性核因子”被鉴定并命名为STAT5,可被IL-2、IL-3、粒细胞巨噬细胞集落刺激因子(GM-CSF)和IL-5等因子激活。STAT6被鉴定并克隆为IL-4诱导因子。
04
JAK-STAT通路信号转导
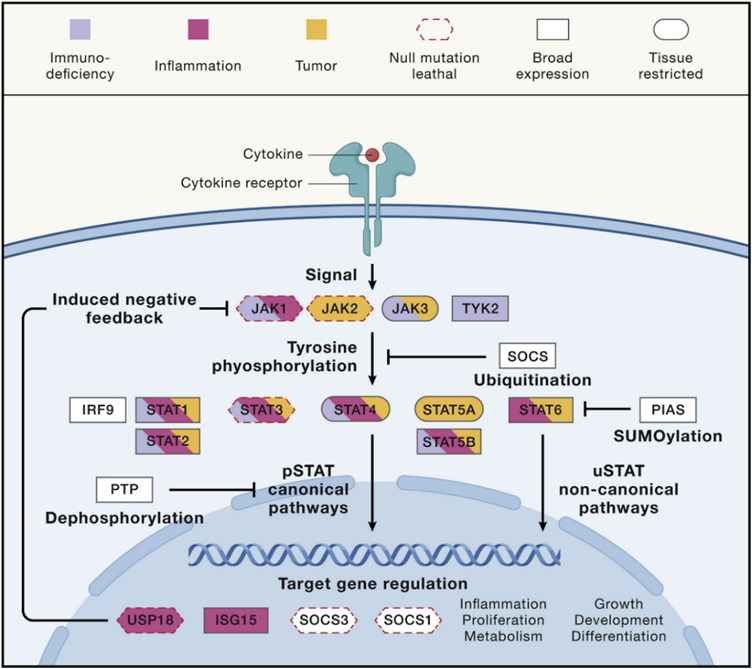
图3 JAK-STAT信号转导
05
细胞因子信号通路的负调控
STAT靶基因的细胞因子信号抑制因子(SOCS家族)的SOCS蛋白通过与细胞因子受体结合,与STATs竞争,干扰细胞因子信号转导。SOCS1和SOCS3的激酶抑制区(KIR)直接与JAK(JAK2、JAK1和TYK2)相互作用。其他关键的负反馈调节因子有泛素特异性肽酶18(USP18)和IFN刺激的基因15(ISG15)。激活STAT蛋白抑制剂(PIAS)家族由多个成员组成,它们作用于STATs和包括NF-kB在内的其他因子,并通过多种机制干扰信号转导。
● 病毒相关的负调控
直接或间接靶向STAT的病毒抗免疫策略突出了STAT介导的IFN信号通路作为抗病毒防御的重要性。STAT导向的IFN拮抗机制包括靶向STAT降解、隔离和细胞质滞留。黄病毒,包括登革热病毒和寨卡病毒,以STAT2为目标进行蛋白酶体依赖性降解。SARS-CoV-2 ORF6蛋白通过Nup98-Rae1核孔亚复合物干扰STAT的输入。SARS-CoV-2 N蛋白也可介导STAT胞质滞留。
06
STAT结构、复合物形成及对功能的影响
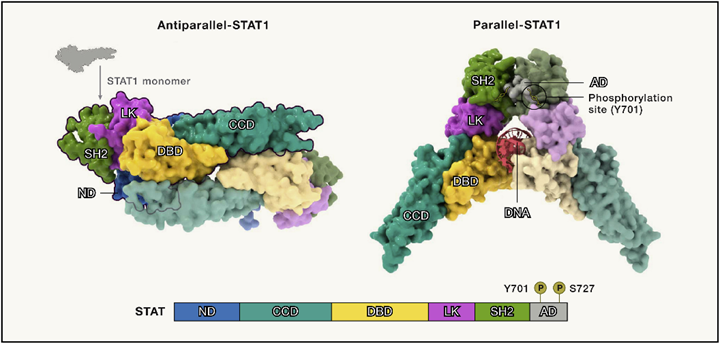
图4 STAT 结构域和结构
● STAT可以形成多种异源或同源二聚体。
STAT1、STAT3和STAT5a已被证明以平行或反平行的构象形成同源二聚体(图4;以STAT1为例)。STAT四聚体通过N端之间的相互作用协同招募同源二聚体成对形成。
响应IFN I和III的基因表达由STAT1:STAT2异源二聚体驱动,并与IRF9一起组成干扰素刺激基因因子3(ISGF3)(图5)。
● U-STAT1:U-STAT2二聚体不仅抑制多种P-STAT1同型二聚体的功能,还抑制ISGF3的形成。复合物形成也影响STAT的核质穿梭。
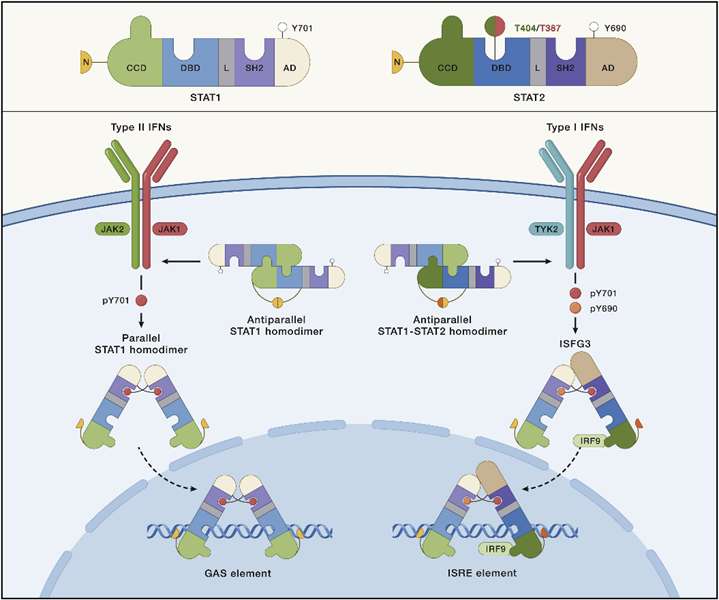
图5 STAT1和STAT2复合物的构象变化
07
STAT和转录调控
●ISG的STAT调节
考虑到对病毒感染的快速反应的必要性,ISG转录的诱导通常是快速的。这与转录速率的增加、快速mRNA加工和降解速率,并且没有转录后控制相对应。RNA Pol II重新招募到ISG是由IFN刺激的,ISGF3结合位点或附近的启动子和基因体的染色质可及性增加,并招募SWI/SNF核小体重塑复合体亚基。组蛋白乙酰化和去乙酰化都是ISG转录激活所必需的。
●STAT和其他转录因子
在生理条件下,细胞接收多个并发信号,激活多个信号通路。STAT和其他TF之间的相互作用包括TF合成的调控、TF活性的调节以及基因组内影响转录的直接或间接相互作用。
●STAT之间的协同作用和拮抗作用
无论信号异同,不同的STAT都可以相互协同或拮抗,从而影响转录。由于 STAT1和STAT3的丝氨酸磷酸化,IL-2和IL-12介导的协同作用,导致更高水平的IFN-γ 表达。STAT之间的拮抗通常在IFN和STAT3激活细胞因子的状态下,这一过程影响多种疾病的发生发展。
08
细胞因子作用的基因组学视图
STAT与基因组的结合有多种影响。干扰素疾病与IFN表观基因组特征相关,因此STAT可以介导诱导和抑制表观基因组和转录组效应。活性增强剂可对STAT产生广泛影响(图6),它们可以影响近端组蛋白修饰,也可以参与潜在增强子的形成,还可以驱动缺乏LDTF结合基序区域的可及性。控制细胞类型特异性功能的关键基因往往受到被TFs、共激活因子、RNA Pol II和延伸因子密集占据的增强子簇的调控。
●STAT可以通过多种机制来传递其作用:直接转录诱导/抑制、表观遗传调节或两者兼有,激活的STAT对表观遗传的调节有重要作用。
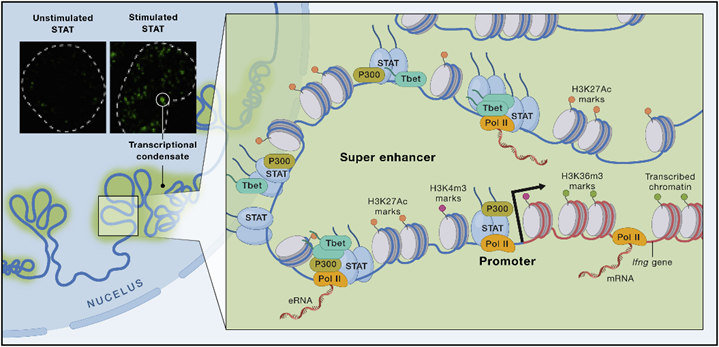
图6 STAT和染色质组织
09
STAT的线粒体定位
10
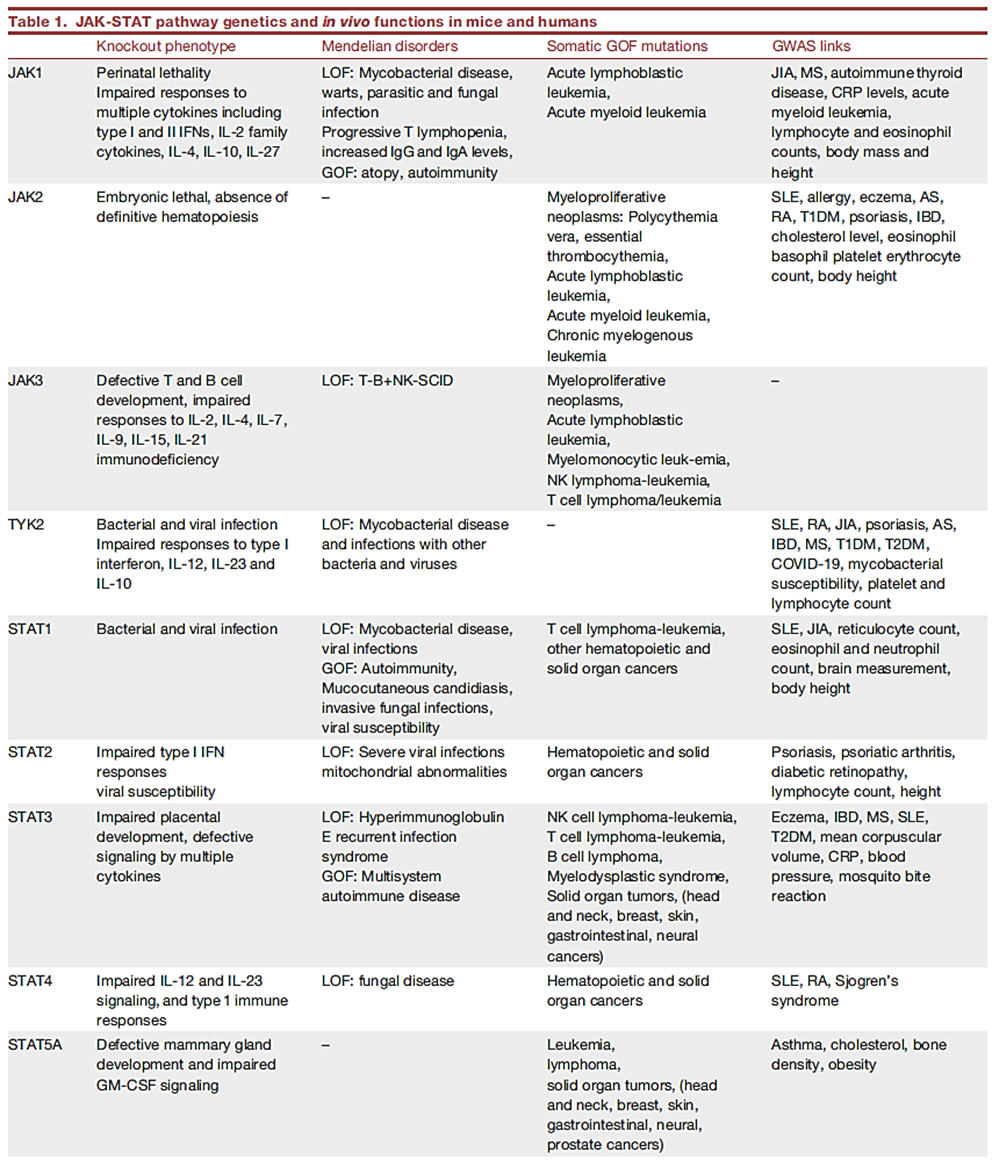
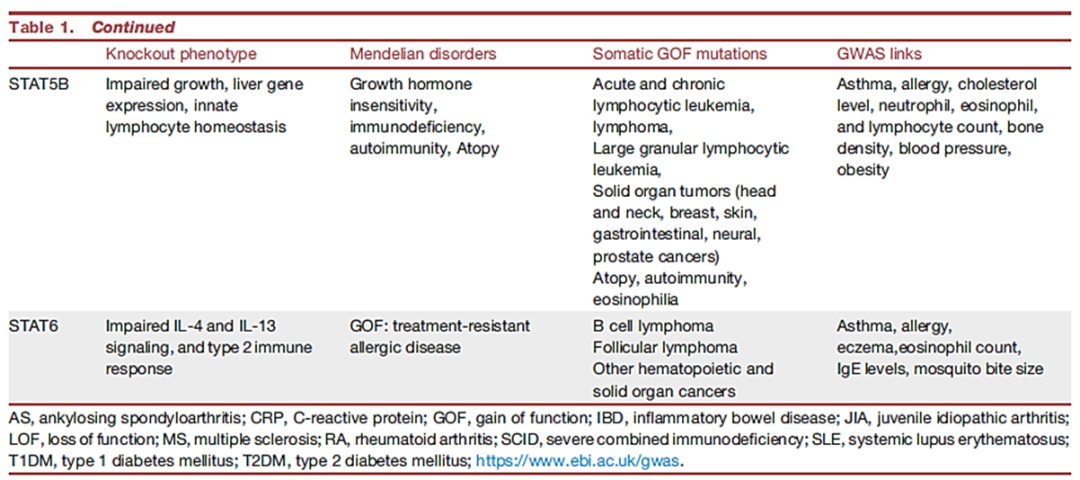
JAKs和STATs的体内功能
1、JAK的关键作用首次在严重联合免疫缺陷(SCID)的人类中被证明。JAK3 常染色体隐性功能丧失(LOF)突变是导致SCID的重要原因。
2、人类TYK2缺乏与细菌和病毒感染以及过敏性疾病有关。人类TYK2的LOF变异(P1104A)与自身免疫性疾病的发病率的降低和结核病风险的增加相关。
3、JAK2对包括造血生长因子在内的多种细胞因子具有重要作用。在骨髓增生性肿瘤(MPNs)患者中发现了JAK2 GOF突变。
4、人类的JAK1 LOF突变与复发性分枝杆菌感染、发育迟缓和早期转移性膀胱癌相关。JAK1 GOF突变导致自身炎症相关疾病和高免疫球蛋白E综合征(HIES)。
●STAT突变
1、STAT1-GOF患者易患自身免疫性疾病,对真菌和病毒感染的易感性增加;
小鼠和人类的STAT2 LOF突变导致了对病毒的易感性增加。
2、STAT3的LOF突变是高免疫球蛋白 E(乔布氏)综合征的分子基础。STAT3 GWAS变异与炎症性肠病 (IBD)、MS、2型糖尿病 (T2DM) 和心肌梗塞有关,具有STAT3 GOF突变的患者好发淋巴造血系统肿瘤和多发性自身免疫性疾病。
3、人类的STAT4 LOF突变与真菌感染和病毒感染的易感性相关。STAT4变异与狼疮、类风湿性关节炎(RA)和干燥综合征相关。
4、人类的STAT5B LOF突变会导致面部畸形和自身免疫。而且还可导致非克隆性高嗜酸性粒细胞增多、湿疹、荨麻疹和腹泻。
5、STAT6 GOF突变与伴有嗜酸性粒细胞增多症的过敏性疾病相关,包括过敏、哮喘和嗜酸性粒细胞增多症。
● 负反馈调节器的突变
小鼠SOCS1缺乏对IFN-γ过敏是致命的,而SOCS1缺陷的人类表现出早发性自身免疫性和过敏性疾病。人类USP18和ISG15缺乏的主要后果是严重的系统性干扰素病。
表1 JAK-STAT 通路体内功能
11
癌细胞中JAK和STAT的复杂角色
肿瘤与GOF突变有关。所有JAK和STAT的突变都很常见,但主要见于造血/淋巴系统恶性肿瘤中。包括乳腺癌、肺癌和结肠癌的非造血系统肿瘤中,JAK和STAT的突变发生较少,但并不意味着该通路与实体肿瘤无关。
在肿瘤微环境和间质癌相关成纤维细胞中,STAT3可以通过改变细胞外基质的产生、血管生成和代谢环境来驱动肿瘤的发展和进展。
12
JAK/STAT通路的治疗靶向性
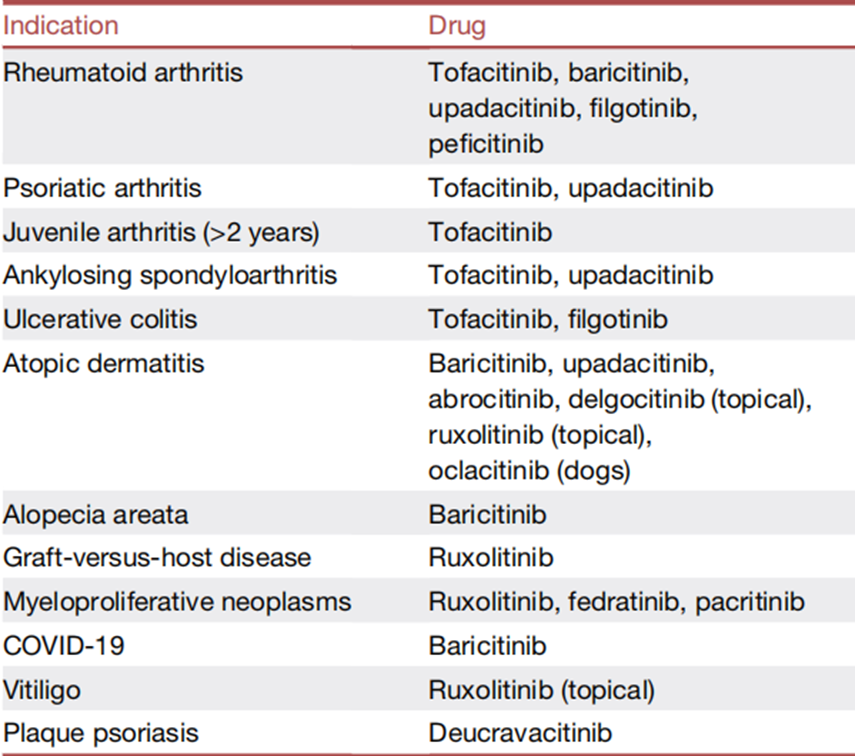
目前11种JAK抑制剂已被批准用于治疗包括RA、银屑病和特应性皮炎等自身免疫系统疾病,移植物抗宿主病,骨髓纤维化,肿瘤,斑秃等各类疾病(表2)。小编之前也整理了截止到今年10月份的药物信息,详见推文:能取代修美乐成为未来药王吗?神奇JAK抑制剂最新汇总
到目前为止,JAK抑制剂的大多数不良事件都可以用已知的特定细胞因子对感染、贫血和癌症的作用机制来解释,其安全性在很大程度上可以与生物制剂相媲美。其他的不良相互作用包括血栓栓塞事件的风险增加,脂质的改变,和心血管并发症。
选择性JAK抑制剂的开发干扰较少细胞因子的功能,能够增加特异性,减少不良反应。
靶向蛋白降解现在也是生成包括JAK1在内的激酶抑制剂的一种选择。
表2 获批的 JAK 抑制剂
●STAT抑制剂
各种策略已被用于开发STAT抑制剂,包括干扰SH2介导的二聚体、DNA结合、或反转录激活和诱导STAT降解。多种抑制剂正在临床试验中,目前进展最快的药物处于临床II期,包括IONIS公司的靶向STAT3反义寡核苷酸Danvatirsen(国内开展,三阴性乳腺癌),Hanlim公司的STAT抑制剂HL-237(韩国开展,类风湿性关节炎),Otsuka公司的STAT抑制剂OPB-111077(新加坡开展,实体瘤)。
●SOCS模拟物
SOCS拟肽药物已在免疫和肿瘤临床前模型中显示出疗效。
13
●结构和细胞生物学
关于细胞因子受体和STAT复合物的结构信息仍然缺乏。随着成像方式的进步,可以使用更复杂的工具来分析细胞因子信号转导的早期事件。
●更全面地理解STAT介导的染色质修饰和转录调控
尽管STAT是信号依赖性TF,但非磷酸化STAT1的作用说明了仍需探究U-STAT1如何修饰染色质和三维基因组。
利用体细胞GOF STAT突变,详细了解STAT对相分离的生物物理方面的结构和功能贡献将对解释增强子在染色质环之外的功能,以及增强子和启动子接近很重要。
●实现细胞、组织和刺激特异性程序
也许最大的挑战是理解细胞如何使用有限的JAK和STAT传递由大量细胞因子产生的信号。
JAK-STAT通路可能有许多属性帮助解决这一生物学难题。
首先是信号成分表达的特异性和调控。
第二,不同STAT激活幅度、磷酸化和去磷酸化速率、核易位和DNA结合不同。STAT异源二聚体的形成被认为是特异性的一种因素。但在没有同源DNA结合基序的情况下,STAT如何被招募仍有待阐明。
第三,许多刺激特异性STAT翻译后修饰状态影响细胞内运输、DNA结合和转录潜力,但这些修饰通常不能全面和动态地测量。但几乎没有关于JAK对不同刺激的翻译后修饰的信息,这需要进一步研究。
最后,多组学工具的产生便于全面测量随着时间变化,在体内单细胞响应生理和相关病理的变化。
对JAK和STAT的更复杂的知识无疑将为具有更大特异性的新药提供许多机会。
Sartorelli V, Horvath CM, Darnell JE Jr, Stark GR, O’Shea JJ. The JAK-STAT
pathway at 30: Much learned, much more to do. Cell. 2022 Oct
13;185(21):3857-3876. doi: 10.1016/j.cell.2022.09.023.
编辑:小果果,转载请注明出处:https://www.cells88.com/linchuang/lcyj/22337.html
免责声明:本站所转载文章来源于其他平台,主要目的在于分享行业相关知识,传递当前最新资讯。图片、文章版权均属于原作者所有,如有侵权,请及时告知,我们会在24小时内删除相关信息。
说明:本站所发布的案例均摘录于文献,仅用于科普干细胞与再生医学相关知识,不作为医疗建议。
 微信扫一扫
微信扫一扫  支付宝扫一扫
支付宝扫一扫 
