

多发性硬化症(MS)是一种中枢神经系统(CNS)的慢性炎症性脱髓鞘疾病,其病程多变且不可预测,可表现为多种生理和认知症状。T细胞和B细胞现在被认为是多发性硬化症发病机制中的中心介质,自从1942年发现CSF-IgG升高以来,B细胞抗体介导的免疫就与MS的发病机制息息相关。
最近,随着以CD20为靶点的B细胞去除疗法的成功,非抗体分泌B细胞作为MS免疫发病机制中的关键角色的作用受到了进一步的关注。这些疗法,包括 rituximab,ocrelizumab 和ofatumumab 减少了新的炎症病变和复发,尽管保留了大多数ASCs,即CD20-浆细胞和一些浆母细胞。在非ASC B细胞亚型中,记忆B细胞(Bmem)在调节MS免疫过程中的作用受到越来越多的关注。Bmem有几个独特的特征,包括延长寿命,对抗原再暴露的快速反应能力,以及作为抗体分泌细胞的前体。
Bmem是免疫记忆中的一个关键角色,它遇到抗原后一直处于静止状态,直到再次暴露于抗原,此时对第二次攻击迅速作出反应。在第一次接触病原体时,大多数Bmem来自生发中心(GC),GCs是次级淋巴组织(SLT)内的特殊结构,在SLT中,成熟的、有抗原暴露的B细胞与T细胞发生同源相互作用,增殖,经历体细胞超突变以增加B细胞受体(BCR)对抗原的亲和力,进行免疫球蛋白(Ig)同型转换。GC-B细胞最终选择分化为抗原特异性、同型转换的抗体分泌(ASC)B细胞或Bmem。
目前,Bmem与ASC分化的调控机制仍不清楚。许多因素已被提出有助于Bmem的形成,动物模型表明,转录因子BACH2选择具有中等亲和力的GC B细胞分化为Bmem。此外,Bmem的产生与ZBTB32、KLF2、ABF-1、STAT5、BCL-6和SKI等因子的表达增加有关,这些因子通常抑制向ASC表型的分化。细胞因子包括IL-24和IL-9能促进Bmem的形成,此外,在体外,IL-2、IL-10和CD40L被证明参与了GC B细胞向Bmem表型的分化。
在体外,Bmem向CXCL12、CCL19和CXCL13迁移,表明这些趋化因子可能参与SLT内的运动和向生存龛或炎症部位的运输。如果长期存在于骨髓中的浆细胞产生的体液免疫不足以消除病原体,Bmem就会积极参与炎症反应。在再次暴露于抗原后,Bmem将产生相对于原始B细胞更快速和有效的抗原特异性反应。
在人类中,Bmem通常通过肿瘤坏死因子超家族成员CD27的表达来鉴定。然而,CD27并非Bmem所独有,CD27也在GC B细胞和后GC B细胞上表达,包括ASC。


因此,CD27的表达应与低水平CD23以及ASC标记物CD138(syndecan-1)的缺乏相结合,以鉴定人类Bmem。进一步包含CD38、CD21、CD24、CD19、B220、FCRL4(FcRH4)以及CD25的特征性标记可以描绘异质性的Bmem群体。
在啮齿动物模型中,Bmem数量少且CD27表达缺失,这阻碍了Bmem的鉴定。小鼠研究已经提出至少10个Bmem亚群,利用Ig等分型结合CD80、PDL-2、CD73、CD38的表面表达。然而,这些标记可能在其他小鼠B细胞亚型上也表达,因此,识别Bmem需要一组不同的表面标记。
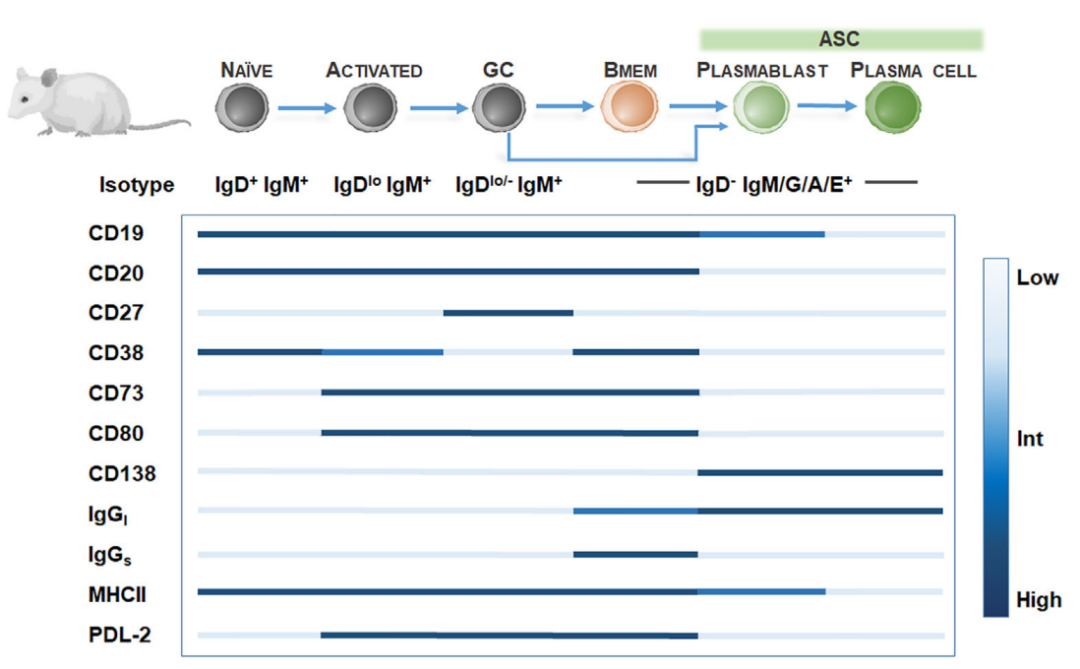
与原始的成熟B细胞相比,Bmem表现出一些独特的特征。Bmem延长了寿命,并且可以不依赖抗原存活数年,甚至伴随宿主的一生。相比之下,原始的成熟B细胞寿命只有几周。与成熟的B细胞不同,Bmem能够快速增殖并分化为ASC,刺激要求很低,包括再次暴露于低水平抗原、T细胞帮助或多克隆刺激。
一旦被激活,Bmem可以遵循两条途径:1)快速分化为ASC;2)重新进入次级GC进行进一步的亲和力成熟和同型转换。Bmem向ASC分化有助于快速、大量地产生高亲和力抗体,以补充终末分化浆细胞产生的抗体。除了向ASC快速分化外,Bmem还是有效的抗原呈递细胞(APC),表达MHCII,不仅能够有效识别抗原,而且能够处理抗原呈递以激活其他免疫细胞,包括T细胞。最后,Bmem可以产生大量的细胞因子,包括TNF、GM-CSF、IL-6、淋巴毒素(LT)和IL-10。
在MS中,B细胞位于中枢神经系统的多个腔室内,包括脑脊液(CSF)、脑实质和脑膜。Bmem在脑脊液高于外周血,并且占到了B细胞的大多数。然而,对MS中Bmem的研究主要集中在外周血和脑脊液。此外,关于Bmem的定义存在显著差异,大多数研究仅基于CD27表达来定义。因此,对于每一项研究,要注意用于定义Bmem的表面标记。
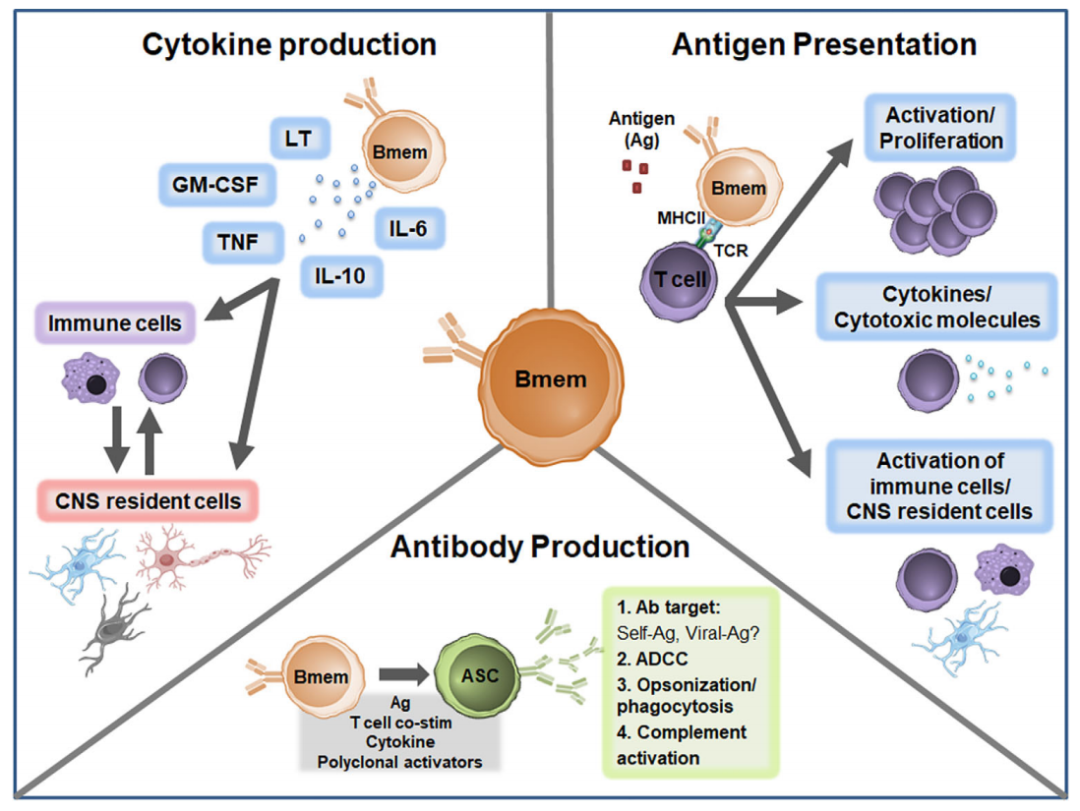
抗体产生和抗原特异性
追踪Bmem转化为ASC以研究体内抗体的产生和特异性仍然具有挑战性,通常需要专门的小鼠模型。或者在体外,利用特异性触发Bmem分化的多克隆激活剂,包括TLR7/8激动剂R848,可以刺激Bmem转化为ASC。随后可量化Bmem,并可评估Ig类型和抗体产生。
Hohmann等人从MS患者的外周血中分离出B细胞,并比较了ASC或Bmem衍生的ASC产生的IgG抗体,研究发现Bmem来源的ASCs分泌IgG能力更强。
MS中的B细胞抗原特异性尚不清楚,但已证明是异质性的,抗体靶点从自身抗原到病毒抗原不等。关于Bmem,目前很少有研究。Hohmann等人检查了对正常人脑裂解物的反应性,在30例复发缓解型多发性硬化(RRMS)患者中,有15例患者的外周血中存在脑反应性Bmem衍生的ASC,脑反应性B细胞(包括Bmem)的存在预示着复发。在健康献血者或其他神经系统疾病对照者的外周血中未观察到脑反应性B细胞。
抗原呈递
Bmem通常被认为是有效的抗原递呈细胞。在MS中,与原始B细胞相比,CSF中的Bmem(CD27+IgD-)显示两种共同刺激分子的表达上调,这两种分子是抗原呈递功能的关键,即CD80和CD86。
与这些发现一致,从RRMS患者分离的离体Bmem(CD19+CD27+)在抗原(包括破伤风类毒素、髓鞘碱性蛋白和髓鞘少突胶质细胞蛋白)存在下能够诱导自体CD4 +T细胞增殖和IFN-γ产生。
细胞因子产生
MS患者的B细胞,包括Bmem,可能表现出一种细胞因子网络失调的倾向。与健康对照组相比,MS患者外周血中Bmem(CD27+)产生GM-CSF增加。此外,与健康对照组相比,从RRMS和SPMS患者外周血分离的体外刺激B细胞显示出抗炎细胞因子IL-10的产生减少,而LT和TNF水平相当。
与原始B细胞相比,从健康捐赠者获得的体外刺激的Bmem也显示出较低水平的IL-10,因此,无论疾病的发病机制如何,低水平的IL-10似乎是典型的Bmem特征。因此,在RRMS和SPMS患者中观察到的B细胞产生IL-10的减少可能归因于另一种B细胞表型,包括产生IL-10的调节性B细胞或ASC。
与临床疾病的关系
在RRMS患者中,与复发期相比,CD5+Bmem亚群的增加与缓解期相关。此外,Nissimov等人证实,外周血Bmem频率升高与扩展残疾状态评分降低相关。相反,Comabella等人确定RRMS患者外周血中非同型转化和同型转化Bmem(CD19+CD27+IgD+或IgD-)的增加与MRI表型高度神经退行性变有关。
儿童和成人MS患者外周血中的Bmem也不同。在小儿多发性硬化症中,与健康儿童和青少年相比,外周血中Bmem(CD20+CD27+)升高。成人MS患者外周血同型转化Bmem(CD20+CD27+IgD-)和浆细胞升高,而儿童MS患者非同型转化Bmem(CD20+CD27+IgD+)和浆细胞增多。
B细胞靶向免疫调节疗法
以CD20为靶点的B细胞耗竭疗法,包括rituximab, ocrelizumab和ofatumumab,清除除了ASC和前B细胞外的所有B细胞,在降低RRMS患者的临床复发率和新病灶形成方面具有显著疗效。抗CD20治疗后,MS患者外周血中包括Bmem在内的B细胞显著减少,治疗后6个月外周血B细胞依然明显减少。
在rituximab治疗的患者中,RRMS患者的脑脊液B细胞也减少了,而PPMS患者的脑脊液B细胞仅显示为中度减少。在RRMS患者中,rituximab治疗可使外周血中产生GM-CSF和IL-10的B细胞比例正常化。治疗后8至24个月,在外周血出现的B细胞中记忆性B细胞明显减少。
进一步的B细胞靶向治疗已经开始寻求针对更多样化的B细胞表型。Inebilizumab(MEDI-551),一种抗CD19单克隆抗体,靶向从前B细胞到浆细胞的所有类型。与同样在CD4+T细胞亚群上表达的CD20不同,CD19仅在B细胞上表达。与抗CD20导向疗法类似,RRMS患者的治疗导致外周血B细胞减少和钆增强病变减少。
针对B细胞生存因子的B细胞免疫调节疗法显示出不同的临床结果。Atacicept治疗RRMS患者导致年化复发率增加,钆增强病变未改变, II期临床试验提前终止。在类风湿性关节炎患者中,atacicept治疗导致外周血中Bmem的数量增加,证实了先前的研究,Bmem可以幸免。类似地,一种能阻断未成熟B细胞、成熟B细胞和ASC存活的抗BAFF单克隆抗体tabalumab也不能清除Bmem。外周血中的Bmem增加,在RRMS患者中未观察到钆增强病变的减少。
最近,以B细胞为靶点的多发性硬化疗法已经扩展到包括布鲁顿酪氨酸激酶(BTK)抑制剂。BTK是通过BCR、FcγR和GM-CSF受体传递信号的关键酶,BTK抑制作用影响骨髓细胞,包括小胶质细胞,以及除T细胞、浆细胞和自然杀伤细胞外的其他造血系细胞。BTK抑制剂evobrutinib、tolebrutinib、fenebrutinib、orelabrutinib和B11091目前正在临床上开发用于复发和进展型MS。在临床试验中,BTK抑制剂可减少钆增强病变和新的或扩大的T2低信号病变,但不能降低RRMS患者的年复发率或疾病进展。初步研究显示,在48周的治疗期间,监测用evobrutinib治疗的RRMS和SPMS患者的外周血B细胞总数或Bmem没有临床相关的变化。
其他免疫调节疗法
许多用于MS的免疫调节疗法也被观察到影响Bmem。尽管传统上意义上认为这些疗法不能调节B细胞,但它们可以直接或间接地影响Bmem的存活和功能。IFN-β、醋酸格拉替雷、芬戈莫德、富马酸二甲酯和米托蒽醌均能减少外周血中Bmem的数量,并改变治疗后的B细胞整体功能。
多种其他免疫调节疗法(包括克拉屈滨、特氟米特、daclizumab和alemtuzumab)均能有效改善RRMS患者的临床转归,这些疗法均能降低外周Bmem数,尽管治疗后B细胞功能改变的相关结果仍有待确定。
MS的病因尚不清楚,但越来越多的证据表明,多种B细胞表型是MS发病机制的核心因素。在多发性硬化症中,外周和中枢神经系统的Bmem正在被越来越多地探索,以确定与疾病发展和进展的确切关系。
已有的研究结果显示,MS患者外周血和中枢神经系统的Bmem发生改变,在抗体产生、抗原呈递和细胞因子产生中发挥潜在的致病作用。利用现有的免疫调节疗法有效地靶向Bmem,可以缓解MS。
未来的研究应致力于解决几个关键的尚未解决的问题,以提供有关MS中Bmem的更深入的见解,包括转运机制、CNS中不同部位内的作用、MS免疫发病机制中的功能相关性,以及确定与临床结果的关联。这些见解可能有助于指导治疗策略,以开发新的药物并改善目前的治疗方案。
参考文献:
1. Memory B Cells in Multiple Sclerosis:Emerging Players in Disease Pathogenesis. FrontImmunol. 2021; 12: 676686.
编辑:小果果,转载请注明出处:https://www.cells88.com/linchuang/lcyj/21087.html
免责声明:本站所转载文章来源于其他平台,主要目的在于分享行业相关知识,传递当前最新资讯。图片、文章版权均属于原作者所有,如有侵权,请及时告知,我们会在24小时内删除相关信息。
说明:本站所发布的案例均摘录于文献,仅用于科普干细胞与再生医学相关知识,不作为医疗建议。
 微信扫一扫
微信扫一扫  支付宝扫一扫
支付宝扫一扫 