

▉ 摘要
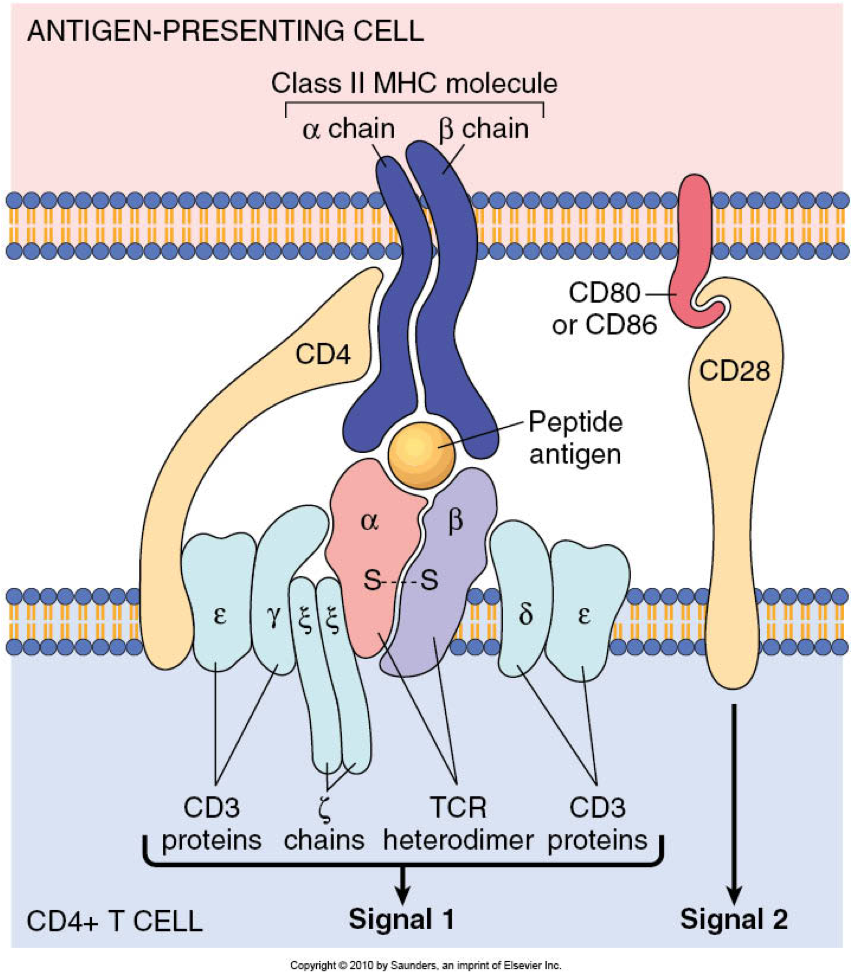

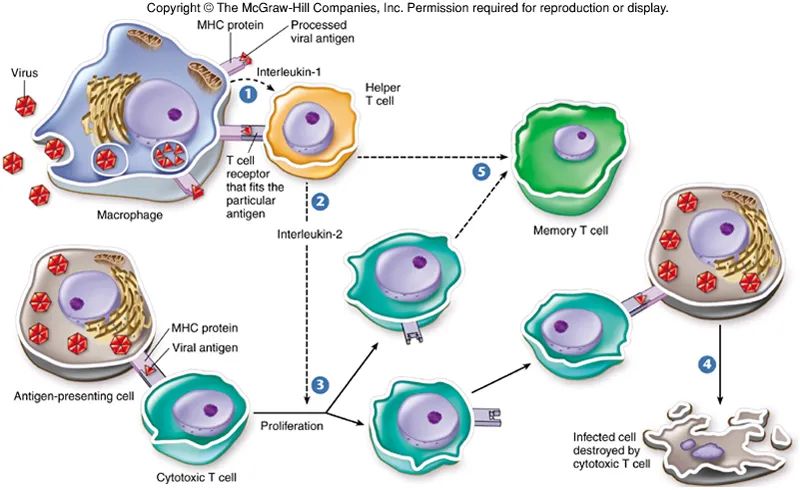
▉1. PI3K-AGC signaling
因此,PI3K活性可以通过将大量含有PH结构域的效应蛋白招募到邻近区域来产生亚细胞信号中枢。PI3K活性诱导参与调节细胞功能的多种信号通路,包括Akt(蛋白激酶B)、磷脂酰肌醇依赖性蛋白激酶1(PDK1)以及mTOR复合物1。
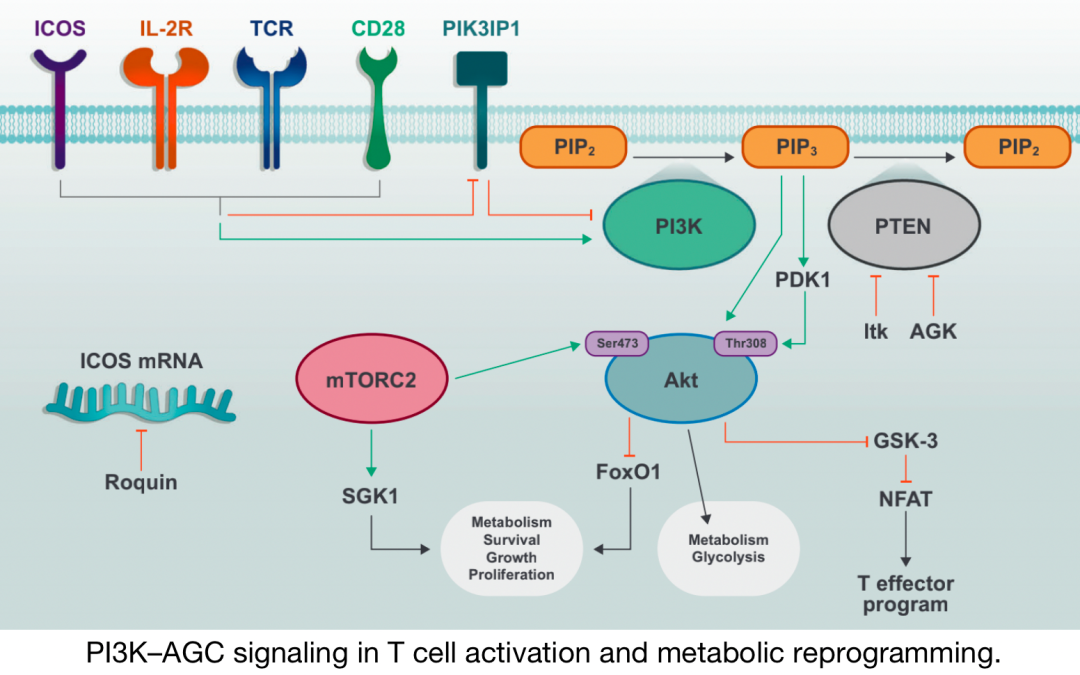
PI3K–T细胞活化和代谢重编程中的AGC信号
PI3K信号由磷酸酶活性负调控。具体而言,PIP3分别通过磷酸酶和张力蛋白同系物(PTEN)以及含SH2结构域的肌醇5′-磷酸酶(SHIP)转化为PIP2或PI-(3,4)-P2。SHIP在促进Th1细胞反应中起着至关重要的作用,而PTEN缺乏会导致T细胞过度活化,尤其是在次优刺激下。PTEN在强TCR刺激或CD28共刺激下被抑制,部分原因是Tec家族激酶IL-2诱导的T细胞激酶(Itk),并且在CD8 T细胞激活期间也受到AGK介导的磷酸化的负调节。
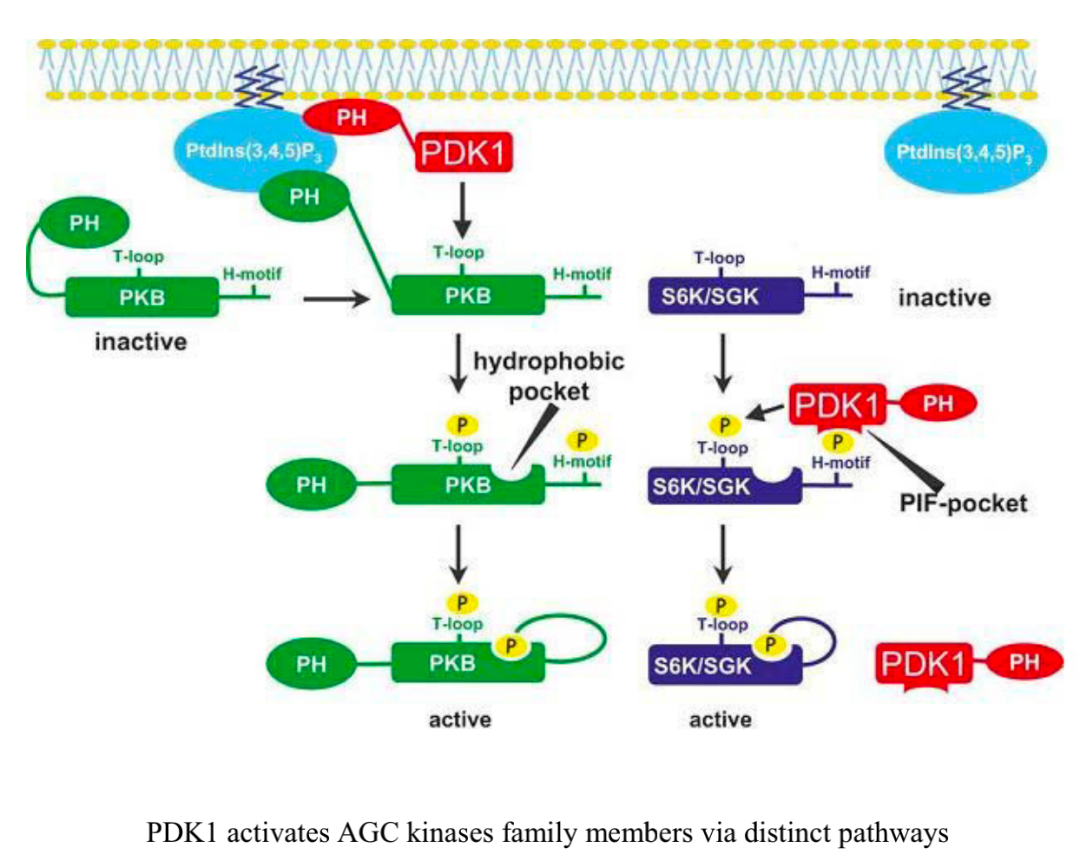
另一种参与Akt信号传导的丝氨酸/苏氨酸激酶是糖原合成酶激酶3(GSK-3),它在静息细胞中具有组成性活性,但通过Akt的磷酸化而失活。活性GSK-3在T和B细胞中通过限制活化T细胞的核因子(NFAT)活性,促进其存活并限制活化。AGC激酶(如RSK)及其底物在T细胞代谢规划中的确切作用一直备受关注,也同样是目前研究的重点方向。
▉2. mTOR signaling
mTOR信号通过调节体内Treg细胞的激活、谱系稳定性和抑制功能,在对抗传统T细胞反应中发挥额外作用。最后,mTOR信号可拮抗淋巴组织中长期记忆CD8 T细胞的分化,却可促进其在非淋巴组织中的发育。因此,mTOR信号是T细胞反应的中央调节器。
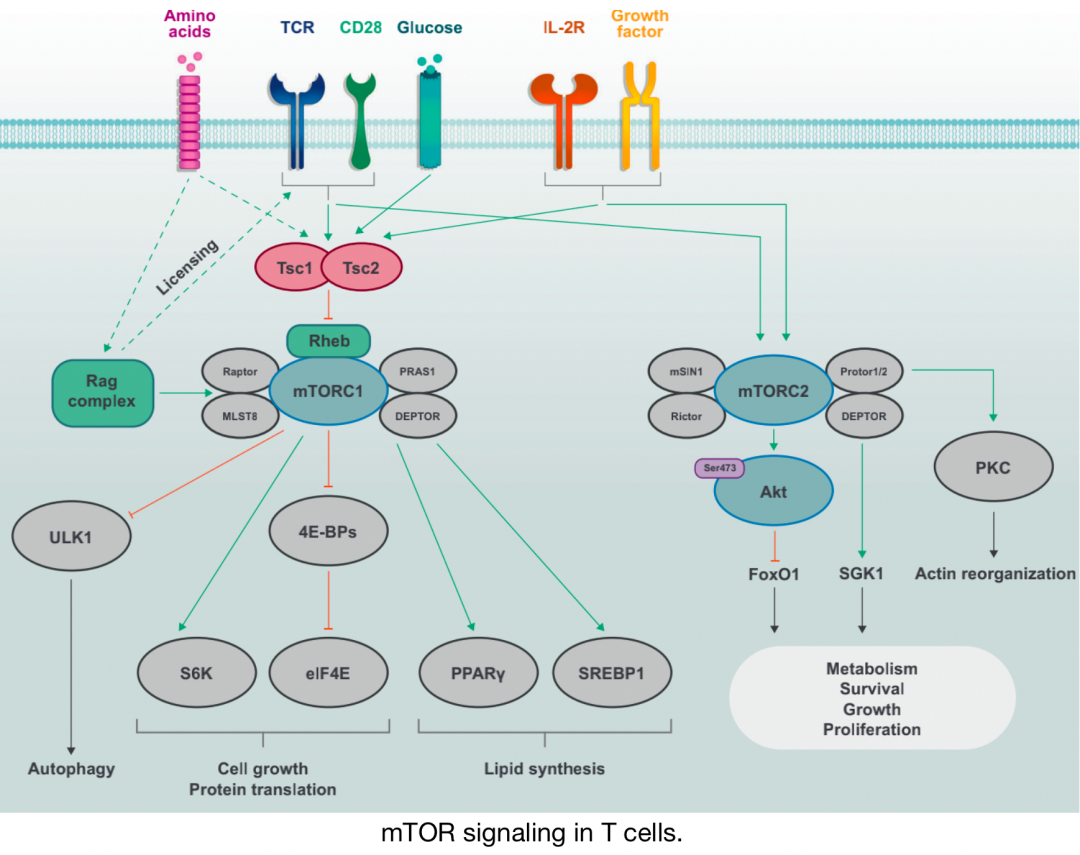
T细胞中的mTOR信号
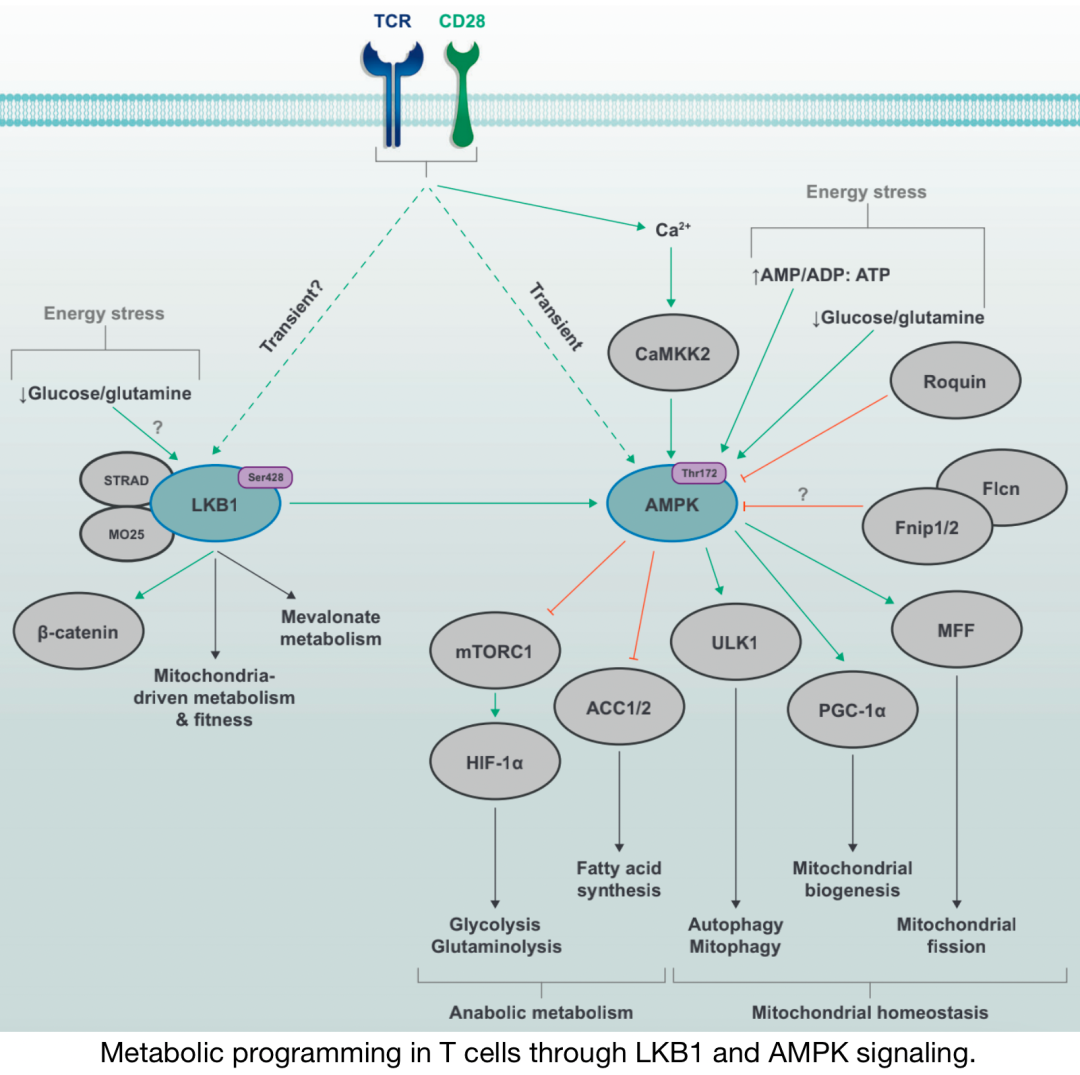
通过LKB1和AMPK信号在T细胞中进行代谢编程
▉3.1 LKB1和AMPK的调节
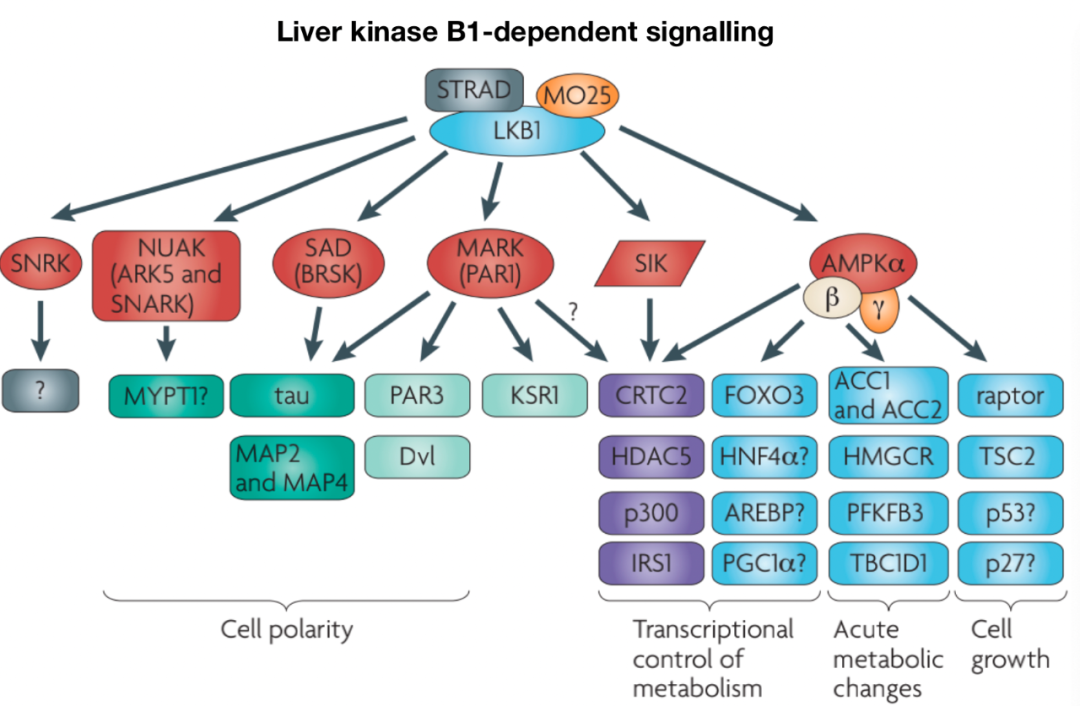
《The LKB1-AMPK pathway: metabolism and growth control in tumour suppression》
AMPK是一种保守的丝氨酸/苏氨酸激酶,由α、β和γ亚基组成。AMP和ADP与CBS3重复序列的结合限制了磷酸酶对AMPK催化α亚基上Thr172的访问,该亚基是促进AMPK激酶活性的关键磷酸化位点。最终,代谢产物的复杂调节允许AMPK活性在不同能量应激条件下的“分级”反应。至少三种上游激酶介导AMPK Thr172磷酸化:LKB1、钙钙调素激酶激酶2(CaMKK2)和TGF-β激活激酶1(TAK1),其中LKB1和CaMKK2的作用最为清楚。在T细胞中,TCR和Ca2信号以及CD28共刺激协同作用,通过CaMKK2快速、瞬时激活AMPK。AMPK活性也通过蛋白质-蛋白质相互作用进行调节。Fnip1和Fnip2与AMPK形成复合物,Fnip1的功能缺失突变提高了B细胞的AMPK活性,从而确立Fnip1是AMPK的负调节因子。Fnip结合蛋白Flcn也与AMPK相互作用,可能作为AMPK信号的负调节因子,有证据表明Fnip1和Flcn受AMPK的相互调节。
▉ 3.2 LKBI-AMPK介导的分解代谢
对肿瘤细胞的研究表明,LKB1或AMPK信号的破坏促进有氧糖酵解,部分是通过HIF-1α、因此增加糖酵解酶的转录。HIF-1α的LKB1–AMPK依赖性调节也可能部分依赖于mTORC1的抑制。LKB1–AMPK信号可能通过HIF-1α或ACC1介导的糖酵解和线粒体氧化代谢变化间接协调Th17和Treg细胞系的分化。此外,最近的研究表明,LKB1促进稳定的Foxp3表达,以及Th2样Treg细胞的发育,独立于AMPK和mTORC1–HIF-1α信号,但依赖于β-连环蛋白信号。LKB1信号是TCR介导的Treg细胞激活后线粒体功能和线粒体依赖性代谢程序所必需的,包括FAO或嘌呤和嘧啶代谢。这些发现强调LKB1和AMPK协调代谢重编程以调节T细胞分化和Treg细胞功能。
▉4. 免疫代谢信号网络
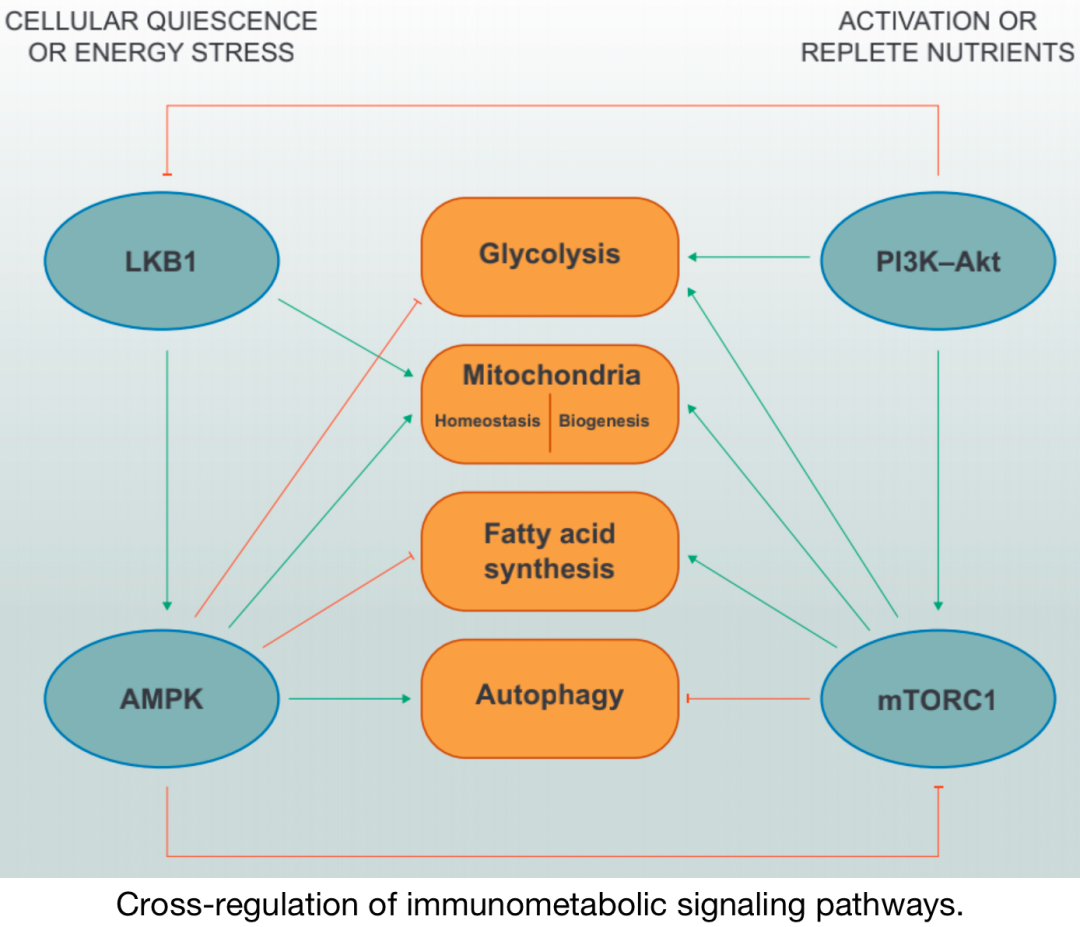
免疫代谢通路的交叉调节
编辑:小果果,转载请注明出处:https://www.cells88.com/linchuang/lcyj/19859.html
免责声明:本站所转载文章来源于其他平台,主要目的在于分享行业相关知识,传递当前最新资讯。图片、文章版权均属于原作者所有,如有侵权,请及时告知,我们会在24小时内删除相关信息。
说明:本站所发布的案例均摘录于文献,仅用于科普干细胞与再生医学相关知识,不作为医疗建议。
 微信扫一扫
微信扫一扫  支付宝扫一扫
支付宝扫一扫 