

介绍
双特异性抗体是具有两种不同类型的结合区域(基于可变结构域的结合位点)的抗体,这使得它们能够同时结合两种不同的抗原。在目前处于临床试验阶段或主要监管机构批准的大约858种抗体中(WR Strohl、BiStro Biotech Consulting Antibody和CAR-T数据库,最后更新于2019年6月20日),目前有122种独特的临床阶段双特异性抗体,其中59它们是CD3ε结合、T细胞重定向双特异性抗体和两种(GT Biopharma GTB-3550和Affimed AFM13,其中是CD16a(FcγRIIIa)NK细胞重定向抗体)(表 2)。


从根本上说,有两种主要类型的双特异性抗体用作TRBA,双特异性抗体片段(例如图 7A-D)和IgG样不对称异双特异性抗体(例如图 7E-H)。这两个主题有很多变化,其中一些对抗体的独特功能至关重要。这些平台的示例如图7所示。有关这些不同平台的更多详细信息可以在最近的各种评论中找到。本节将简要介绍其中一些平台,以及结构如何对TRBA的功能非常重要。
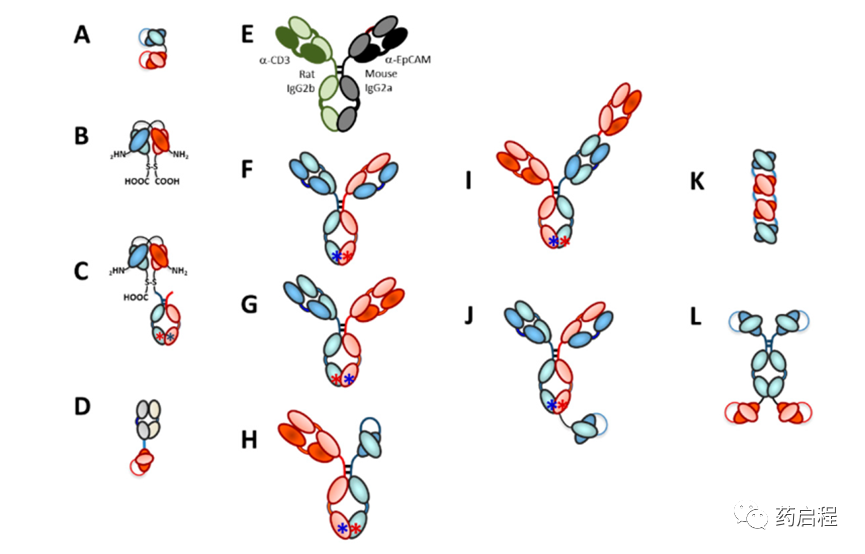
用于制造TRBA的双特异性二价抗体片段
今天开发的大多数双特异性抗体片段都以某种方式基于scFv抗体,由两个研究小组在1988年独立发现,它们由一个VH结构域通过一个短而灵活的接头连接到一个VL结构域组成。值得注意的是,休斯顿等人利用(GGGGS)3柔性连接器将VH融合到 VL;这种“gly-ser”接头或其变体,是当今双特异性抗体中使用最广泛的接头之一。
从历史的角度和治疗的角度来看,最重要的双特异性抗体片段类型是串联 scFvs、双特异性 T 细胞接合剂(BiTE®s[166])、双亲和重定位(DART®s[153]) ) 抗体、双抗体和串联结构域抗体。
最近描述的其他基于片段的双特异性抗体构建体,例如HSA抗体片段融合 、双特异性杀手接合器(BiKE)、三特异性杀手接合器(TriKE)和对接锁定Fab也被开发为双特异性抗体片段平台。
如前所述,抗 CD19×抗CD3 BiTE®blinatumomab已被批准用于商业用途,商品名为Blincyto®。包括 blinatumomab,目前有27个二价双特异性抗体片段在临床试验中作为TRBA进行测试。其中,11个是短半衰期BiTE®或类似于BiTE®的基于双特异性scFv的分子,6个是下一代半衰期延长的BiTE®(即 BiTE®-Fcs),一个是短半衰期BiTE®半衰期 DART® 构建体,三个是长半衰期DART®-Fc分子,三个是抗癌免疫动员单克隆TCR (ImmTACs) ,一个是三特异性杀手接合剂(TriKE) ,两个是三特异性T细胞激活构建体(TriTACs)。
用于制备TRBA的双特异性二价不对称IgG样抗体
当今制造用作TRBA的双特异性抗体的最广泛使用的方法之一是产生含有两种不同类型重链的不对称异双特异性IgG。制造不对称IgG样双特异性抗体需要两个重要成分,这些抗体可以以一致的方式开发和制造。首先是不对称异二聚重链(HC)的形成和/或分离优于亲本IgG同二聚抗体。第二个是每个臂的轻链(LC)与同源HC的正确配对。
1 HC的不对称配对
众所周知,HC二聚化的主要驱动因素是CH3-CH3结构域之间的高亲和力(约10 pM)相互作用。CH3-CH3结构域之间的相互作用,其表面积超过 2400Å2,由一个强大的中央疏水核心驱动,该核心被一系列带电残基包围,提供两个 Fc 结构域之间的静电相互作用。
本质上,已使用三种基本策略来促进不对称异二聚化而不是亲本同二聚体的形成:(i)第一种基于物理/空间相互作用,例如,在一条重链上添加一个突出物,在另一个重链上添加相应的裂隙(例如,“旋钮入孔”);(ii) 第二个取决于 CH3-CH3 界面处特定氨基酸相互作用的改变和(iii)第三个关注电荷,即改变带电荷的氨基酸残基以产生同二聚体的排斥和相应的电荷异二聚体对之间的吸引力 。
几种不同的平台已被用于制造用作TRBA的不对称异源双特异性IgG样抗体。第一个是小鼠B细胞、大鼠B细胞和骨髓瘤细胞的三向融合,形成四瘤细胞系(Triomab® 技术)(图 7E)。在这个平台中,由于物种特异性,LCs自然地分类到适当的重链。当然,缺点是使用这种方法制备的任何抗体都将具有高度免疫原性,如前所述,这是catumaxomab的问题之一。该平台还用于生成第一个不对称异二聚体IgG样TRBA,于1995年进行临床测试。
所有其余的不对称双特异性IgG样平台都依赖于Fc的工程改造,以促进异二聚 IgG 的形成或纯化,而不是亲本同二聚IgG。第一个真正的双特异性IgG样抗体平台是旋钮入孔(KIH)平台。其他依赖电荷吸引/排斥的平台包括静电转向(ES)平台、ES 加铰链突变、基于Oncomed IgG2的ES“Bimab”技术和Chugai“Asymmetric Re-工程技术免疫球蛋白”(Art-Ig®)平台。
其他不对称异双特异性IgG方法包括修饰CH3结构域界面中的特定氨基酸配对,例如Duobody®方法、主要基于疏水相互作用的Zymeworks azymetric平台、BEAT(通过Glenmark、Xencor H/A平台和Merus Biclonics® 平台的基于T细胞受体的抗体)平台。用于制造不对称异源双特异性IgG样抗体的其他平台包括Regeneron 的修饰蛋白A结合平台,即“链交换工程域”(SEED),它由衍生自IgG和IgA CH3域的交替序列组成,导致不对称但互补对,AG和GA,只有异二聚体蛋白才能结合并折叠成活性Fc和NovImmune的kλ抗体平台,该平台在一个Fab臂中包含与λ LC配对的常见HC和kappa LC在另一个Fab臂中。
在这种情况下,基本上所有的结合活性都取决于LC ,这与使用具有普通LC的异源HC的情况完全相反。
2 不对称异双特异性IgG样抗体的LC问题
无论是通过融合两个不同杂交瘤还是通过基因工程产生杂交杂交瘤,将编码两条不同重链(HC)和两条不同轻链(LC)的四种抗体基因引入细胞系,将导致形成由于混杂的重链(HC)-轻链(LC)配对,十种不同的潜在组合中只有一种是所需的双特异性抗体。如果两条不同的重链强制配对以形成不对称的异源Fc,则这种LC配对问题可以减少到四种可能的配对。因此,即使通过产生不对称Fc高效形成异二聚体Fc,仍然需要解决轻链独立分布问题。
已发现多种解决方案可缓解不对称双特异性IgG样抗体中的轻链配对问题。一种方法,本质上是一种“避免”策略,是形成不对称的IgG样抗体,一侧有Fab臂,另一半有scFv(图 7H),例如Xencor的Xmab H/A平台或Glenmark的BEAT平台 。第二种解决方案是“通用 LC”方法,其中不对称IgG样双特异性抗体的两个Fab都具有相同的LC。另一种策略是通过切换一半抗体上的CH和CL结构域来产生HC-LC相互作用的差异,以产生“CrossMab”(CM)。
这种转换的结果是双特异性抗体一侧的一条正常重链和轻链配对,另一半的 VH-CL-hinge-CH2-CH3与VL-CH1配对(CMCH1-CL;)。与CrossMab类似的策略是在一个和/或另一个Fab臂中突变HC和LC界面中的某些序列,以确保正确配对。
控制LC分布的最后一种方法是通过两个亲本抗体的单独上游生产与两个抗体半的后蛋白A重组,通常通过还原和再氧化过程,导致不对称异二聚体双特异性抗体。围绕这个概念构建了两个平台,Duobody®平台和SEEDbody平台。这些方法取决于这样一个事实,即重链可以通过减少链间二硫键来分离,而HC-LC相互作用和二硫键保持稳定。这个主题的一个变体是在两个共培养的大肠杆菌菌株中产生双特异性抗体,每个菌株含有半抗体 (HC+LC)。在菌株生长和抗体半体产生后,抗体被还原并重新氧化以形成异二聚体IgG 。还设计了一种用于产生不对称异二聚体双特异性抗体的哺乳动物共培养方案,结合了Duobody®和细菌方法的各个方面。
除了在欧洲被批准用于恶性腹水但现已停产的 catumaxomab 之外,目前有 21 种已知的不对称二价异双特异性TRBA正在使用上述设计的临床试验中,所有这些都旨在通过CD33ε结合重定向 T 细胞,杀死目标细胞(表2)。
3 三价双特异性抗体平台
最近开发了几个平台,为靶细胞提供两个结合臂,为CD3ε提供一个结合臂,从而产生三价但双特异性的抗体。这些抗体形式背后的概念是通过亲和力更好地结合靶细胞,同时只为CD3ε提供一个结合臂,因为众所周知,为CD3ε提供两个结合臂可能会导致非特异性和不希望的T细胞活化。
一种三价双特异性抗体形式是不对称异二聚体IgG,使用KIH技术构建,如前所述,单个抗CD3ε Fab臂附加到 HC 之一(图 7I)。通过使用CrossMab技术来维持LC保真度。这些2:1(靶细胞抗原:CD3ε)TRBA被罗氏称为“TCB”,即“T细胞双特异性”。目前有两个临床阶段的 TCB,将 Fab 作为额外的附件,包括 cibisatamab(又名 RG7802、RO6958688、CEA TCB)[159,204],它有两个用于 CEA 的结合臂和一个用于CD3ε和RG6026的单个结合臂(又名 RO7082859 ),它有两个用于 CD20 的绑定臂和一个用于 CD3ε的绑定臂。已经报道了另外两种TCB,一种靶向 BCMA,另一种靶向在大约一半HER2阳性肿瘤中表达的HER2的羧基末端片段 ,但尚未进行临床试验。
另一种三价双特异性抗体是ERY974,它是一种沉默的IgG1不对称mAb,其抗 CD3 scFv与其中一条重链的C 端序列融合(图 7J)。ERY974具有两个glypican-3 结合位点以优化亲和力,但只有一个抗CD3臂可减少非特异性T细胞激活的机会,目前处于 I 期临床试验。
其他为靶细胞提供两个结合位点和为T细胞CD3ε提供单个结合位点的三价双特异性抗体平台是“用于T细胞激活和癌症杀伤的不对称串联三聚体”(ATTACK)和新的三价IgG 形抗体平台tri-Fab 格式。这些平台尚未在诊所中出现。
4 四价双特异性抗体平台
通常认为CD3ε的二价靶向可导致非特异性T细胞激活和细胞因子释放,这在T细胞重定向平台中是不可取的。因此,如前几节所述,绝大多数片段和IgG样TRBA都是二价的,一个抗原结合位点与靶细胞结合,另一个抗原结合位点与 T 细胞上的CD3ε结合。
然而,尽管如此,一些临床阶段的平台脱颖而出,成为这一趋势的对立面。其中第一个是Affimed串联双抗体(TandAb)平台,它是一种双特异性、四价串联双抗体,分子量约为114kDa。临床阶段的TandAbs包括AMV564,Aphivena用于骨髓增生异常综合征的抗CD33×CD3 TRBA和AFM13,Affimed的NK细胞重定向抗CD30×CD16a 候选药物用于霍奇金淋巴瘤。
Aptevo ADAPTIRTM平台是一种四价双特异性抗体,由两个与靶标A结合并与 Fc 铰链融合的相同scFv和两个与靶标B结合的相同 scFv 通过短接头融合至该 Fc 的 C 端序列组成(参见图 7L) 。抗前列腺特异性膜抗原 (PSMA)×抗CD3四价双特异性ADAPTIRTM TRBA 分子 MOR209(也称为 ES414)目前正在 1 期临床试验中进行潜在治疗前列腺癌的测试。在临床前研究中,尽管 MOR209 拥有两个CD3ε 结合区域,但MOR209似乎不会不加选择地激活T细胞,并且它诱导的促炎细胞因子水平低于其他一些平台。因此,TRBA 几何形状可能在与T细胞表面上的两个 CD3ε 结合是否导致非特异性 T 细胞活化方面发挥重要作用。
目前在临床试验中未出现的其他四价双特异性平台包括IgG-scFv融合、双可变域免疫球蛋白(DVD-Ig) 和Fabs-in-tandem免疫球蛋白 (FIT-Ig)。IgG-scFv 融合体是 IgGs,scFvs融合到每个HC或LC的C端或N端 ,由于 scFv 的展开和聚集而经常遭受不稳定性,需要额外的修饰以实现稳定,可制造的候选。DVD-Igs是约200 kDa的四价双特异性抗体,由IgG组成,在N端附加一个额外的Fv。然而,开发DVD-Ig的Abbvie团队已将其TRBA结构转移到半DVD平台,以减少非特异性T细胞激活。最后,Epimab Biotherapeutics “Fabs-in-tandem immunoglobulins” (FIT-Ig)平台在某些方面与DVD-Ig相似,不同之处在于每 HC的N端都附加了一个完整的 Fab。目前临床上没有来自这些平台的 TRBA。
影响TRBA效力的因素
影响TRBA效力的因素包括靶抗原上表位的位置、靶抗原的大小、TRBA臂对靶抗原和CD3ε的亲和力、TRBA对靶抗原的化合价、抗体大小和几何形状、抗原密度靶细胞、效应物与靶标的比率和TRBA浓度。江等人使用多种参数建立靶细胞-生物-效应细胞复合物 (TBE复合物) 模型,以证明多种参数对杀伤效力的敏感性。出现的一个关键结果是TRBA浓度似乎很关键,TRBA浓度大于低亲和力结合臂的KD,由于向单价结合转变,导致TBE复合物形成减少。另一方面,过高浓度的TRBA会导致抗原阳性细胞和CD3ε阳性 T 细胞分别被覆,从而导致杀伤效果较差。
虽然影响TRBA效力的因素的层次结构尚未完全了解,但在过去几年中,一些因素已经变得清晰。首先,抗体与靶抗原结合的表位至关重要,最好的表位是膜近端,尤其是对于较大的靶抗原。研究表明,靶向膜近端表位也会导致突触中负调节蛋白CD45的排除,从而增加 T 细胞反应和杀伤的效力。基于TRBA的癌症靶点P-钙粘蛋白和ROR1研究支持了靶抗原上的膜近端表位应提供最大TRBA效力的这一观察结果。其次,抗原的大小可以有效地增加T细胞和靶细胞之间突触内的距离,也可以影响效力,更大的目标会导致更低的TRBA效力。
已经多次表明,增加 TRBA 对靶抗原的亲和力可以显着提高效力。增加TRBA与靶细胞结合的另一种方法是增加亲合力,即在靶细胞上具有更多的结合臂。有两种TRBA格式为靶细胞提供两个结合臂,只有一个 CD3ε 结合臂,罗氏科学家设计的三 Fab TCB格式和ART-Ig ®-scFv格式由 Chugai 科学家设计。
了解 2:1 构建体是否比1:1构建体显示出更好的效力的关键在于为同一目标制造每种类型的构建体。巴卡克等人比较了他们的2:1抗CD20×CD3ε与类似制造的1:1构建体,并证明 2:1格式在体外的效力比类似的1:1构建体高10-100倍。此外,2:1格式在离体试验中优于1:1格式,并在体内动物模型中显示出非常有效的活性。此外,Bacac等人证明,在用抗CD20×CD3ε 2:1 TCB、RG6026治疗之前,用抗CD20 mAb、obinutuzumab进行预处理,导致细胞因子释放显着降低,这可能转化为临床益处。
考虑到抗CD20×CD3ε 2:1 TCB格式似乎比各种 1:1 抗 CD20×CD3ε格式更有效,看看其他多肿瘤靶点结合×单CD3ε 绑定格式可能会起作用。为此,IGM Biosciences 在临床前研究中拥有一种抗 CD20 × CD3ε IgM五聚体抗体,该抗体具有10个CD20结合臂和一个CD3ε结合臂。关注这种高度狂热的抗体(在目标端),看看它最终与1:1和2:1格式的比较会很有趣。
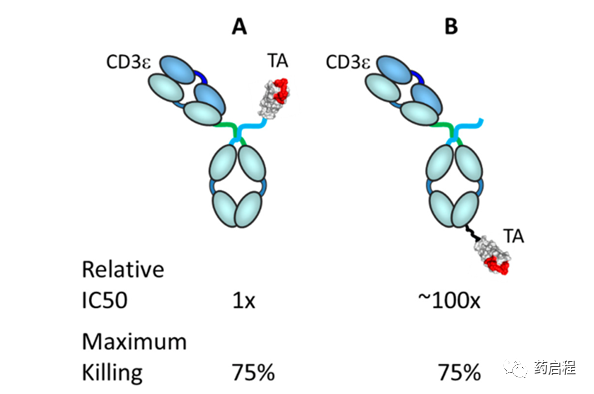
值得注意的是,抗体形式和大小,从小抗体如BiTE®s (ca.24 kDa) 到更大的形式如不对称IgGs(ca150 kDa),似乎对效力的差异影响低于距离膜的表位或对靶抗原的亲和力。此外,结合几何结构(包括靶表位、抗体大小和形式、结合角度以及可能的其他局部因素)会影响TRBA的效力。在Janssen R&D进行的一项未发表的研究中,制备了几种版本的双特异性抗体(CD3ε臂)/centyrin(肿瘤抗原臂)组合,并在体外测试了肿瘤细胞杀伤活性。如图8所示,肿瘤抗原结合臂在分子上的位置,即CD3ε臂与肿瘤抗原臂之间结合的几何形状/距离,约为体外杀伤活性相差 100倍。
另一个关于 T细胞重定向尚未完全研究的领域是Fc功能的作用。设计 Removab®的Triomab® 平台具有高活性Fc结构域,可与人类FcγR相互作用以增强免疫反应。人们普遍认为,T细胞重定向双特异性抗体中活性Fc的存在会增加肿瘤微环境中T细胞和其他效应细胞释放促炎细胞因子的可能性。这些促炎细胞因子的释放被认为是这些抗体的治疗作用机制的一部分,因此虽然这是理想的,但也需要加以控制。另一方面,目前大多数片段-Fc、不对称IgG或附加IgG平台都使用了静音或沉默的Fc,以免通过与骨髓效应细胞的相互作用过度刺激免疫系统。即使没有Fc活性,许多使用T细胞重定向双特异性抗体的治疗也伴随着细胞因子释放综合征(CRS),需要作为治疗范式的一部分加以解决。
因此,现在和将来制造的许多(如果不是大多数)T细胞重定向抗体似乎很可能会继续避免Fc活性以限制免疫介导毒性的可能性。
Bispecific T-Cell Redirection versus Chimeric Antigen
Receptor (CAR)-T Cells as Approaches to Kill Cancer Cells
Antibodies 2019, 8, 41; doi:10.3390/antib8030041
编辑:小果果,转载请注明出处:https://www.cells88.com/cells/myxb/10490.html
免责声明:本站所转载文章来源于其他平台,主要目的在于分享行业相关知识,传递当前最新资讯。图片、文章版权均属于原作者所有,如有侵权,请及时告知,我们会在24小时内删除相关信息。
说明:本站所发布的案例均摘录于文献,仅用于科普干细胞与再生医学相关知识,不作为医疗建议。
 微信扫一扫
微信扫一扫  支付宝扫一扫
支付宝扫一扫 