

原文标题Enhancement of therapeutic potential of mesenchymal stem cell-derived extracellular vesicles,作者Kyong-Su Park等来自瑞典哥德堡大学萨尔格伦斯卡医学院研究所Krefting研究中心。
在对间充质干细胞 (MSCs) 在细胞治疗中的应用进行临床研究之后,人们对其分泌的可溶性因子的兴趣日益增加。随着对 MSCs 及其分泌因子的研究,MSCs 释放的细胞外囊泡 (EVs) 已成为一种新的细胞间通讯工具和串扰模式。
MSC 衍生的 EV 已被确定为在生理和病理条件下必不可少的信号传导介质,它们似乎对 MSC 的许多治疗效果负责。在几个体外和体内模型中,已观察到 EV 在调节免疫系统方面具有支持性功能,主要由 EV 相关蛋白和核酸介导。此外,已证明用生物物理或生化信号(包括来自其他细胞的 EV)刺激 MSC 会影响后续 MSC 衍生的 EV 的内容和生物活性。
该综述概述了 MSC 衍生的 EV 的内容及其支持作用,并就操纵 MSC 以改善 EV 的分泌和随后的 EV 介导的活动提供了不同的观点。在这篇综述中,研究人员讨论了制备MSCs 进行基于 EV 的细胞治疗以及使用 EV 影响 MSCs 中的元素表达的可能性。
通过这种方式,人们对 EV 在以 MSC 为重点的细胞治疗中的最新进展提供了清晰的视角,并提出了相关的问题和建议,以填补知识空白。
细胞外囊泡 (EVs) 与外泌体(Exosomes)
细胞外囊泡 (EVs) 是由细胞释放的强大生物实体,其中包含可以促进其靶点变化的分子。因此,EV 已被研究用于临床应用,如疫苗、免疫抑制剂或修复和分化过程的刺激剂。
EV 是一个总称,包括各种不同的释放囊泡,例如外泌体和微泡 (MV)。“外泌体”通常用于描述源自内体起源的多泡体与质膜融合的囊泡。这种生物发生将它们与其他 EV 区分开来,例如,通过质膜出芽释放的那些,通常称为 MV。由于它们独特的生物发生机制,MV 通常比外泌体大(通常,外泌体的直径小于 200 纳米,而 MV 的尺寸范围可达 1000 纳米),但这些尺寸范围可能会重叠。
EV不同的分离方案侧重于分离富含外泌体、MV 或两者的组分。由于细胞外囊泡的研究是一个全新的领域,许多研究在报告细胞外囊泡的数据时使用了异构命名法。通常会找到诸如“外泌体”、“MV”和“微粒”之类的术语,指的是模糊的 EV。为了统一起见,研究人员在这里将所有这些研究都包括在“细胞外囊泡”一词下。
尽管许多功能都归因于EV,尤其是参与细胞通讯,但它们在体内的作用仍然知之甚少。可能仍有一些主要功能和作用仍然未知,不同细胞在病理状态下释放的 EVs 的免疫学作用仍缺乏研究。
另一方面,由于已知分子对这些囊泡的分类和 EV 释放的模式反映了它们的起源细胞类型和生理状态,因此细胞外液的 EV 分数可以提供非常丰富的信息。因此,大量注意力集中在使用含有来自受伤组织和肿瘤的 EV 的“液体活检”来检测疾病生物标志物,以期开发具有高灵敏度和特异性的侵入性较小的诊断程序。将分子分类到 EV 中仍然是一个有点模糊的过程,但它显然涉及不同蛋白质和核酸的富集。作为改变靶细胞中基因和蛋白质表达的试剂,EV 的蛋白质和 RNA 含量受到了特别关注 。
目前,对干细胞分泌囊泡的关注最广泛地涉及间充质干细胞(MSCs),也称为间充质基质细胞。这些细胞是多能细胞,可以从各种成人组织中分离出来。研究最多的 MSCs 是从骨髓 (BM-MSCs)、脂肪组织 (AD-MSCs) 或脐带 (UC-MSCs) 中分离出来的。分离的 MSC 通常是异质的,含有干细胞、定向祖细胞和分化细胞。
尽管没有针对 MSCs 的特异性标志物,但它们通常具有在体外化学诱导后分化成至少三种中胚层细胞谱系(成骨细胞、成软骨细胞和脂肪细胞)的能力 以及不存在造血谱系标记,但存在表面相关标记,如 CD44 和 CD90。
间充质干细胞通过培育和促进周围细胞的增殖和分化来支持它们在体内的生态环境。当移植用于细胞治疗时,这些细胞会迁移到炎症和损伤部位,并且以其在各种疾病模型中促进免疫调节和组织修复的能力而闻名。然而,在没有支架的情况下,它们通常不会永久植入受伤组织,因此它们只会影响目标组织。
MSCs 及其囊泡的分泌组特别令人感兴趣,因为这些细胞主要用于细胞治疗,因为它们具有旁分泌/内分泌作用,而不是它们的分化潜能。除了这些细胞分泌物中存在的可溶性因子(如生长因子和细胞因子)外,MSC 培养物的上清液还富含 EV。许多临床前数据的例子表明,源自 MSCs 的 EVs 继承了其原始细胞的治疗效果,使用 EVs 代替细胞本身可以具有以下优势:
绕过与细胞疗法有关的大多数安全问题,例如致癌细胞的细胞污染和不受控制的细胞分裂;
-
能够对这些纳米颗粒进行广泛的潜在操作,以改善递送和应用效果;
-
促进开发优化 MSC 使用的方法,以获得更高产量和更佳的治疗产品。
间充质干细胞对环境变化也非常敏感,在体外不同刺激下表现出不同的分泌谱系和表型,这可能与它们在体内对不同炎症或损伤环境的反应具有很大的动力学有关。
用源自其他细胞(如肥大细胞和上皮细胞)的 EV 处理 MSC 会影响它们的表型,用可溶性因子处理和细胞培养条件的变化也会影响它们的表型。对于科学界来说,更好地控制 MSCs 的免疫调节和分化能力,以设计更有效和更具体的治疗策略,无论是直接细胞治疗还是 EV 介导的治疗,这将是非常有趣的。
MSC 衍生的 EV 已成为各种动物模型中免疫调节和再生效应的有吸引力的介质。基于 EV 的方法已被认为是一种安全且有吸引力的治疗干预措施,但一个显着的限制是 EV 的产量通常较低。
为了克服这个问题,一些高通量自动化程序已应用于大规模细胞和细胞外囊泡生产。最近的研究利用由脂肪干细胞和肿瘤细胞通过连续挤压产生的模拟 EV 纳米囊泡,以克服自然产生的 EV 低产量问题。此外,在许多先前和正在进行的研究中,各种生物物理和生化信号已被证明有助于 EV 的治疗效果并提高其生产水平。
基于 EV 的治疗也面临着关于 EV 制剂纯度的挑战。我们使用术语“MSCs + 细胞外囊泡 + 外泌体 + 微囊泡”的 PubMed 文献搜索确定了几种不同的 EV 分离方法(图 1)。超过一半的文章仅使用超速离心,约 27% 使用商业试剂盒,主要基于蛋白质沉淀方案。只有大约 19% 的文章使用了某种方法将游离分泌蛋白与 EV 分离(例如,密度梯度、过滤和阴离子交换)。为了使我们对 EV 衍生功能的常识保持一致,有必要将 MSC 衍生 EV 的真实成分与分泌蛋白分开研究。生产高纯度的 MSC-EV 制剂将加速基础研究结果转化为临床实践。
图1 几种不同的 EV 分离方法
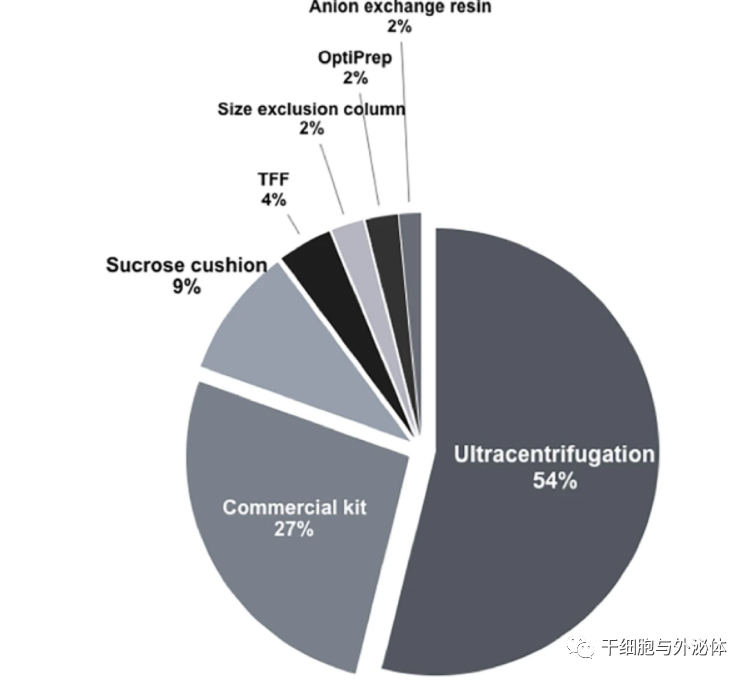

| 分离 MSC-EV 的实验方法,目前共有126篇文章用于分析隔离方法,大多数研究是使用通过超速离心分离的 EV 和 ExoQuick-TC试剂盒等商业套件进行的,TFF,切向流过滤。
在这篇综述中,研究人员讨论了用于治疗应用的 MSC 衍生的细胞外囊泡 (MSC-EV) 的开发。首先,他们将讨论 MSC-EV 的成分及其在不同体内和体外模型中的作用,然后将讨论培养 MSC 以改善或改变其分泌 EV 从而提高其治疗潜力的一些可能性.
MSC-EV 的治疗效力在哪里?
间充质干细胞通过直接的细胞间串扰以及通过分泌广泛的可溶性因子发挥其在体内的作用。MSCs 分泌的主要可溶性介质包括细胞因子、生长因子和 miRNA,它们具有从肿瘤调节、免疫抑制和血管生成到组织再生的多种治疗作用。
最近,除了这些可溶性因子之外,一些研究小组已经开始在 MSCs 的条件培养基 (CM) 中发现另一种功能成分。研究表明通过超速离心分离的 MSC-CM 可抑制小鼠急性肾小管损伤,这种颗粒状部分包括纳米级的囊泡结构。
另一组利用通过 HPLC 衍生的过滤排除获得的 EV 部分,其中包括带有 EV 标记蛋白的囊泡,以减少急性心肌梗塞的大小,这在之前使用 MSC 和可溶性因子的研究中已经完成。
除了上述使用 MSC-CM 识别具有治疗功能的 EV 的研究外,还有大约 126 篇已发表的文章讨论了 EV 在各种疾病模型中的治疗功能。在这里,我们将重点介绍已被证明具有不同功能效应的 MSC-EV 相关货物(蛋白质和核酸)(图 2、表 1 和附加文件 1:表 S1)。
图 2 MSC 衍生 EV 的成分及其相关的治疗潜力
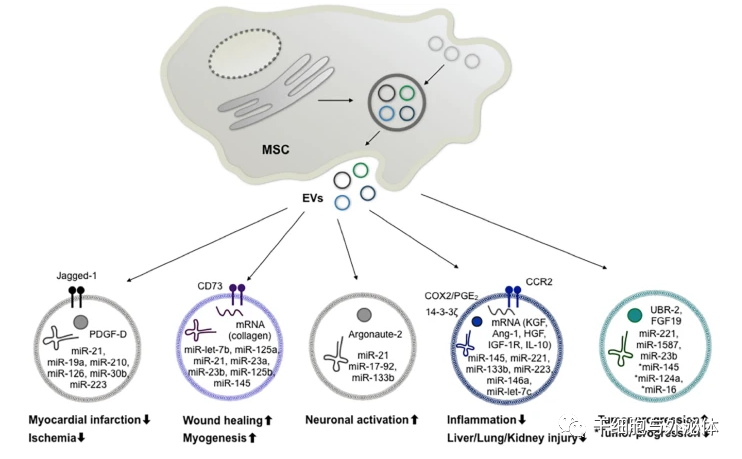
| 不同的圆圈显示了介导 MSC-EV 的免疫调节、再生和肿瘤生长作用的建议的泡状蛋白、mRNA 和 miRNA 成分。缩写:MSC,间充质干细胞;EVs,细胞外囊泡;PDGF-D,血小板衍生生长因子-D;COX2,环氧合酶 2;PGE2,前列腺素 E2;CCR2,C-C 趋化因子受体 2 型;KGF,角质形成细胞生长因子;Ang-1,血管生成素-1;HGF,肝细胞生长因子;IGF-1R,胰岛素样生长因子 1 受体;IL-10,白细胞介素 10;UBR2,泛素蛋白连接酶E3组分n-识别素2;FGF19,成纤维细胞生长因子19。
表 1 在动物模型和体外针对各种疾病进行的 MSC-EV 相关研究
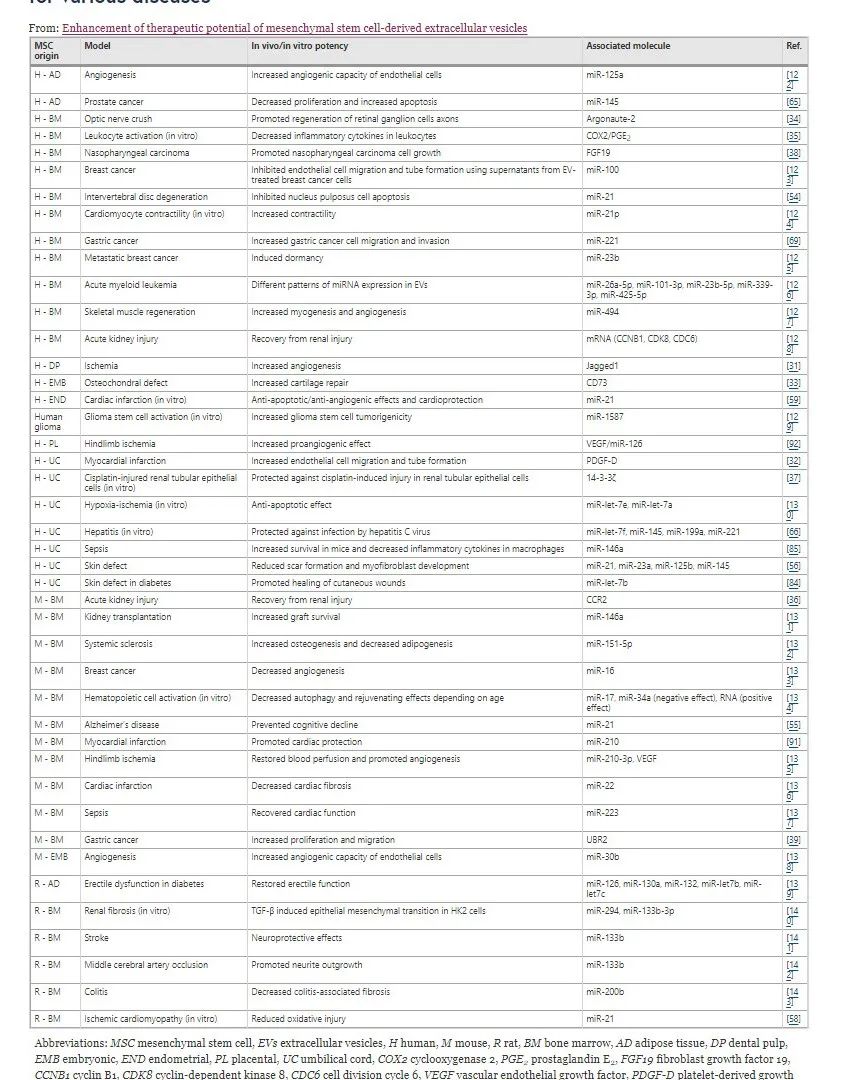
MSC-EV 内的蛋白质效应器
EVs 通常包括完整的膜蛋白,如四跨膜蛋白、外周膜蛋白和胞质蛋白,并且 EVs 蛋白质组成的变化已被证明与重要的功能变化有关。MSC-EVs 还含有许多蛋白质成分,这些蛋白质成分被认为与许多疾病的康复有关。
囊泡蛋白效应物已被探索为通过促进血管生成来治疗缺血和心肌梗塞。例如,来自牙髓来源的 MSCs 的 EV 含有 Jagged-1 配体蛋白,它是 Notch 信号的激活剂,它们被证明可有效激活血管生成信号。
含有 Jagged-1 的 EV 会触发内皮细胞中 Notch 靶基因的转录变化,导致诱导血管生成和毛细血管样管信息,而这种血管生成作用可以用抗 Jagged-1 抗体阻断。
除此之外,UC-MSC-EVs 已被证明携带血小板衍生生长因子-D (PDGF-D),已被证明可有效协助梗塞心脏细胞的组织修复功能。从转染 PDGF-D-siRNA 的 MSCs 中分离出的 EVs 消除了恢复,因此表明 PDGF-D/PDGF 受体相互作用可能在 EV 介导的心肌修复中起关键作用。
在骨再生方面,2018年新加坡国立大学博士Zhang Shipin等人证明了囊泡 CD73 的治疗效果,其中来自胚胎干细胞衍生的 MSC 的 EV 上存在的 CD73 能够修复软骨细胞培养物中的骨软骨缺损,同时巨噬细胞浸润更多一种抗炎表型。使用 CD73 抑制剂的 Akt 和细胞外信号相关激酶 (Erk) 信号传导证实了 CD73 在 EV 中的作用。
此外,还进行了一项神经元再生研究,以研究 BM-MSC-EVs 治疗外伤性和退行性眼病的效果。结果表明,携带 argonaute-2 (AGO-2) 蛋白的 EV 促进了视网膜神经节细胞的显着存活和轴突的再生。在敲除 AGO-2 后,来自 MSCs 的 EV 减弱了这种影响,这表明 AGO-2 参与了 EV 的再生作用。
基于 MSCs 众所周知的免疫调节作用,MSC-EVs 也被描述为抗炎剂,从而使 EVs 用于治疗包括肾损伤在内的免疫疾病合理化。美国德克萨斯大学麦戈文医学院的MT Harting等人研究表明 BM-MSC-EVs 中环氧合酶 2 和前列腺素 E2 的表达增加,这些成分部分导致脾细胞中促炎细胞因子的减弱。
此外,促炎细胞因子 CCL2 通过其存在于 BM-MSC-EVs 上的受体的淬灭作用导致巨噬细胞活化减少并有助于修复急性肾损伤 。此外,通过 EV 递送 14-3-3ζ 可防止通常由化疗药物顺铂引起的自噬小管上皮细胞损伤。
有趣的是,MSC 衍生的 EV 不仅在可用于治疗的组织修复和抗炎作用方面有益,而且癌细胞可以有效地利用 MSC 的功能来实现自身的生长和免疫逃逸。例如,BM-MSC-EV 中存在的成纤维细胞生长因子 19 可促进鼻咽癌细胞生长。同样,BM-MSC-EVs 可以传递泛素蛋白连接酶 E3 组分 n-识别素 2,对胃癌细胞具有增殖和迁移作用。
总体而言,MSCs 的蛋白质货物可以通过淬灭一些促炎因子或通过增强抗炎因子直接发挥功能作用。其中一些影响可能是与其他货物的组合影响,因此将这些组件逐一剖析是设计更有效的 MSC-EV 的一种方式。
MSC-EV 内的核酸
a.NDA)(脱氧核糖核酸)
在 EV 中可以找到多种形式的核酸,包括 DNA、mRNA 和 miRNA。EVs中DNA的存在和定位仍然存在争议,没有研究表明DNA参与MSC-EVs的治疗效果。有趣的是,尽管描述了存在于其他来源的囊泡中的外源 DNA 的促炎作用以及 MSC 对它们的摄取,但我们没有发现任何由 MSC-EV 相关 DNA 诱导的炎症报告,这表明这些EV 的免疫抑制特性可能会克服这种可能的影响,或者与 MSC-EV 相关的有害 DNA 较少。
b.mRNA
与一长串研究相比,很少有研究将治疗效果归因于 mRNA,这些研究至少显示特定 miRNA 与观察到的结果之间的相关性,如表 1 所示。EV 中核酸的化学计量和每种 miRNA 的最小浓度在受体细胞中促进强大作用所需的 mRNA 或 mRNA 仍然是一个深入研究的主题。
在急性肺损伤模型和肺炎中,角质形成细胞生长因子 (KGF) 的 mRNA 与 MSC-EV 治疗观察到的免疫调节有关。在这些研究中,抗 KGF 中和抗体的施用以及治疗消除了最初观察到的对存活的有益作用,并且用针对 KGF 转录物的 siRNA 预处理 MSCs 也部分抑制了 MSC-EVs 的抗炎作用,如支气管肺泡所证明的那样。灌洗液细胞结构和炎性细胞因子的存在。
作者进一步假设血管生成素-1(在 MSC-EV 中也很丰富)的转录本通过在体外和小鼠模型中使用 MSC-EV 在恢复肺蛋白通透性和解决炎症方面发挥重要作用急性肺损。事实上,在这些模型中,MSCs 或 MSC-EVs 的 angiopoietin-1 siRNA 预处理导致人肺微血管内皮细胞的免疫调节和通透性恢复降低。
在急性肾损伤的体外模型中,有研究人员提出了肝细胞生长因子 mRNA 的特殊作用,因为用 RNase 处理的囊泡在促进肾小管细胞的去分化和随后的生长方面无效。在顺铂诱导的急性肾损伤的另一个体外模型中,发现胰岛素样生长因子 1 受体的 EV 相关 mRNA 对于保护近端肾小管上皮细胞很重要。在类似的顺铂诱导的体外模型中,还发现白细胞介素 (IL)-10 mRNA 通过 MSC-EV 转移。
此外,发现用于合成 VII 型胶原的 mRNA 与胶原蛋白本身一起在体外转移到隐性营养不良性大疱性表皮松解细胞中。这种情况的特点是 VII 型胶原基因的功能丧失突变,因此 MSC-EV 可能是治疗这种疾病的潜在方法。
c. miRNA
越来越多的证据表明 MSC-EV 中包含的 miRNA 的有效性。图 2 和表 1 显示了许多参与 MSC-EV 在不同疾病条件下的治疗效果的 miRNA。
由于 miRNA 领域在癌症相关研究中得到了最广泛的探索,因此已知其中一些 miRNA 被上调或被认为是特定癌症类型的标志物。然而,这并不一定意味着 EV 中这些 miRNA 的存在代表了促肿瘤风险,因为它通常是多种因素的组合,对于确定每个分子在该过程中的最终作用很重要。然而,重要的是要记住,当体内缺乏抑癌基因时,致癌分子可能会通过 EV 转移并可能影响肿瘤的发展。另一方面,这种转移在非突变细胞中可能只有短暂的影响。
一般而言,在最常与 MSC-EV 的治疗特性相关的 miRNA 中,miR-21、miR-19a 和 miR-210 与心血管疾病有关;miR-let-7b、miR-125a 和 miR-21 与伤口愈合有关;miR-21、miR-17-92 和 miR-133b 与神经损伤有关;miR-223、miR-146a 和 miR-let-7c 与保护肝脏和肾脏损伤有关;miR-221、miR-1587 和 miR-23b 与癌症相关效应有关(图 2)。在这里,我们将深入讨论一些最常被认为是 MSC-EV 效应的可能介质的 miRNA。
- miR-21
鉴于已显示 miR-21 通过刺激增殖和抑制不同细胞类型的细胞凋亡来调节细胞存活 ,这种 miRNA 的贡献与 MSC-EV 介导的各种疾病模型中的治疗效果有关。BM-MSCs 已被证明可通过 EV 传递外源性 miR-21,从而防止髓核细胞凋亡并减少椎间盘退变。此外,已显示 miR-21 在缺氧条件下在 MSC-EV 中的表达增加,注射这些 MSC-EV 可以减少小鼠的认知和记忆障碍,同时减少斑块沉积和小胶质细胞的活化。
研究人员进一步描述了 miR-21 的功能,来自富含 miR-21 的 UC-MSC 的 EV 在抑制肌成纤维细胞形成从而防止过度瘢痕形成方面发挥关键作用。在这些 EV 中阻断 miR-21 会消除 EV 抑制肌成纤维细胞形成的能力,这表明这种特定的 miRNA 对于 MSC 的抗瘢痕形成功能至关重要。
miR-21 也被描述为在心脏损伤中具有保护作用。源自 BM-MSCs 的 EV 在过氧化氢诱导的氧化后具有增加的 miR-21 水平,并且囊泡 miR-21 可以被转运至心脏干细胞,从而在功能上抑制磷酸酶和张力蛋白同源物 (PTEN) 的表达,从而防止氧化压力触发的细胞死亡。
另一项研究表明,通过抗 miR 治疗对 miR-21 的选择性拮抗消除了 MSC-EV 的抗凋亡和血管生成作用,随后上调了 miR-21 靶标 PTEN,这表明 miR-21 可能是MSC-EVs 对心血管疾病的治疗作用。
- miR-145
miR-145 与细胞分化过程以及平滑肌细胞和肌成纤维细胞的激活有关。此外,miR-145 通常被描述为具有肿瘤抑制作用。与这些发现一致,MSC-EV 中 miR-145 的上调已被证明对皮肤缺损愈合有效,并且在前列腺癌中具有抗肿瘤作用。
miR-145 在 UC-MSC(脐带来源间充质干细胞) 衍生的 EV 中富集,这是通过高通量 RNA 测序确定的。EVs 中 miR-145 的过表达可以抑制肿瘤生长因子 (TGF)-β/SMAD2 的激活,从而抑制成纤维细胞向肌成纤维细胞的分化,而这种 miRNA 的消耗极大地消除了 EVs 抑制 TGF-β 的能力/SMAD2 通路。
在癌症预防方面,AD-MSC(脂肪组织来源的间充质干细胞)衍生的 EVs 通过细胞凋亡显着抑制转移性前列腺癌的增殖,这种作用被 miR-145 敲低所抵消,导致 Caspase 3/7 的表达减少和抗凋亡蛋白的表达增加。有趣的是,UC-MSC 分泌的 EV 已被证明通过抑制病毒感染来抑制丙型肝炎病毒 (HCV) 感染,这主要归因于 miR-145 抑制病毒 RNA 复制[66]。
- miR-221
与 miR-145 相比,miR-221 在癌症进展中的促进作用近年来已得到广泛认可。例如,肝细胞癌中的 CD44 表达由 miR-221 通过 PI3K-Akt-mTOR 通路控制。此外,miR-221 可以通过靶向金属肽酶 2 的组织抑制剂来支持非小细胞肺癌。
同样,已证明 miR-221 在 BM-MSC(骨髓来源的间充质干细胞) 的 EV 中的高表达可有效增加胃癌细胞迁移、侵袭和与细胞外基质的粘附。另一项使用 miR-221 的研究表明,MSC-EV 中上调的 miR-221 以与上述 miR-145 相似的方式保护免受 HCV。
如何使基于 MSC 的疗法更有效?
生物物理信号
MSCs 已被证明受到各种不同的生物物理和生化信号的刺激(图 3)。生物物理诱导剂包括电脉冲、低功率激光照射、非相干红光、电磁场暴露、机械信号(例如,流体、张力和压力)和底物形貌和刚度、2D 和 3D 支架/无支架培养和磁力。
通过这些不同的治疗,MSCs 可能会显着改变其表型并开始分化成特定类型的细胞,这对于组织再生等一系列应用非常有用,尤其是在对间充质起源器官的损伤中。
然而,MSC 培养的生物物理参数中的一些变化也会影响它们的分泌谱,而不会促进完全分化。例如,这些治疗中的许多可以增加 MSCs 的增殖,但人们对它们对 EV 分泌或免疫调节能力的影响知之甚少,留下了广泛的条件有待探索,以试图增加 MSC-EV 产量和控制他们的内容。
图3:各种条件对细胞外囊泡生产和功能的影响
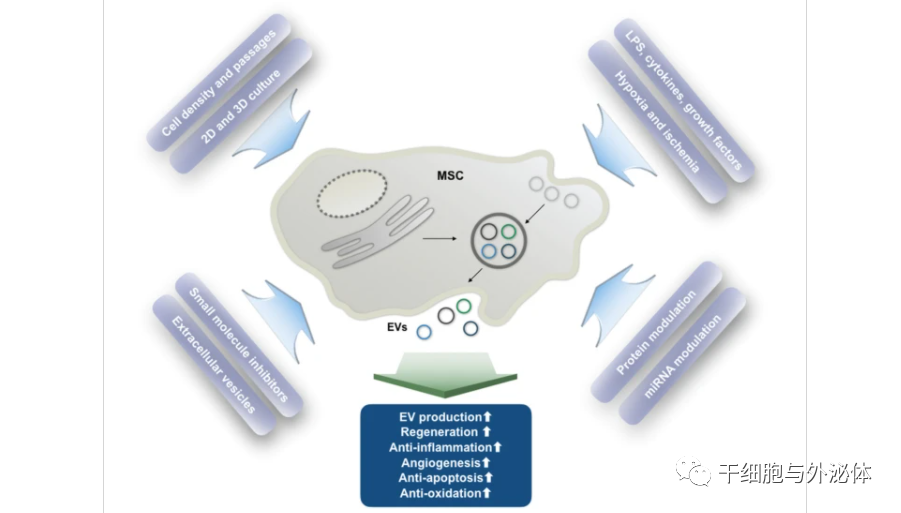
| 培养条件和外部刺激对 MSC-EV 分泌谱和功能变化的影响。MSC,间充质干细胞;EVs,细胞外囊泡;LPS,脂多糖。
在一项针对 I 型胶原支架中 3D 培养的 MSC 与普通 2D 培养的 EV 的研究中,作者使用商业试剂盒从其上清液中分离出 EV,发现在促进功能恢复和免疫调节方面有更多的蛋白质和更好的结果。从 3D 培养细胞中分离的样本中的创伤性脑损伤模型。
另一个可以影响 EV 产量的 MSC 培养参数是细胞接种密度,较低的密度与较高的产量有关。然而,尚不清楚这些影响是否与细胞间接触有关,因为在同一时间段内多次收集而不是一次收集也会增加可收集的 EV 数量。细胞培养物中存在的 EV 或代谢物有可能在生化上降低 MSC 的 EV 产生和/或分泌。
生物化学信号
人们认为,MSCs 对脂多糖等细菌衍生分子和响应这些分子释放的细胞因子有反应,可以提高其对炎症环境的治疗效果。最近,在炎症条件下由 MSC 产生的 EV 变得越来越重要。
一项研究已经表明,脂多糖刺激增加了 UC-MSCs 的 EV 分泌,并增强了 M2 巨噬细胞极化和糖尿病皮肤伤口愈合。越来越多的证据表明,炎性细胞因子可能会增强 MSC-EV 的治疗效果。
来自 IL-1β 预处理的 UC-MSCs 的 EV 显示出比来自未经处理的 MSCs 的 EV 具有更大的免疫调节作用,这表明更多的功能分子(如 miR-146a)嵌入到来自 IL-1β 预处理的 MSCs 的 EV 中。与此一致,在存在肿瘤坏死因子 α、干扰素 γ 或 TGF-β 的情况下培养的 MSC-EV 导致脾细胞中的细胞因子表达显着降低,并强烈增加调节性 T 细胞分化,进而发挥抗炎作用。
MSC-CM包括各种生长因子,如血管内皮生长因子和PDGF,它模拟了与完整细胞相关的有益作用。研究人员利用AD-MSC-EVs的血管生成潜力进行再生医学的工作基础上,证实了用PDGF预刺激诱导的治疗效果增加。
另外,与未经处理的EVs相比,用红细胞生成素的激素刺激增加了EVs的产生,并增强了EVs在肾脏损伤后的保护作用。此外,缺氧和缺血条件已被证明可以改变间充质干细胞在EV功能方面的特征。
据报道,缺氧预处理使BM-MSCs增加EVs的产生,并且这些EVs通过刺激新生血管而在心脏保护方面有卓越的活性。另外,在一氧化氮刺激期间,间充质干细胞释放的EVs已被证明能增强内皮细胞的血管生成作用,并能恢复后肢缺血的肢体功能。
此外,将紫杉醇纳入BM-MSC-EVs被证明可以在体外抑制肿瘤的生长。此外,培养基的血清含量被发现可以改变MSC的特性和释放的EVs的RNA含量,这表明MSC-EVs可以被调控,以包含不同的活性成分用于未来的治疗应用。
来自分化细胞的 EVs 能够改变 MSCs 的特性。来自神经元细胞的 EV 可通过 miR-125b 转移介导 MSC 神经元诱导,而内皮细胞衍生的 EV 影响 MSC 增殖和迁移,为 EV 作为内皮细胞和 MSC 之间的通信通道提供了证据 。
此外,肥大细胞衍生的 EV 通过囊泡相关的 TGF-β 调节 MSC 功能,在卵白蛋白诱导的过敏模型中诱导抗炎作用。源自肿瘤细胞的 EV 也可以调节 MSC 表型。例如,来自癌症干细胞的 EVs 诱导 MSCs 中的化学吸引增加,导致肿瘤进展,而来自肺癌细胞的 EVs 刺激 MSCs 中 IL-6、IL-8 和单核细胞趋化蛋白 1 的产生和分泌,从而灌输 MSCs具有更多的肿瘤支持特征。
然而,尽管这些来自分化细胞的 EV 很可能能够调节 MSC 进一步产生 EV,但就 MSC-EV 而言,其他类型的 EV 如何影响 MSC 功能还没有明确的研究。因此,需要研究其他细胞衍生的 EV 在 MSC 分泌 EV 方面的作用,以获得最佳治疗效果。
MSC的细胞重编程
尽管 MSC-EVs 具有很强的治疗作用,但仍需要进一步了解 MSCs 的基因修饰如何提高分泌型 EVs 的治疗效力。研究人员目前正试图通过蛋白质和 miRNA 的过表达来开发治疗上更优化的 MSC。在这里,作者将重点关注转基因 MSC-EV 如何显示改变的货物和改善的功能效果。
a. MSCs中过表达蛋白
大多数在 MSCs 中过表达的蛋白质都是转录因子和信号分子。MSC 通常具有有限的扩展能力,因此 研究人员通过诱导 c-Myc 的过表达创造了永生化的 MSC。
在严格的 GMP 条件下,从这些永生化细胞生产 EV 是可扩展的,这使得这些 EV 能够用于临床。另一项研究表明,BM-MSC 中 GATA-4 转录因子的过表达增加了其分泌的 EV 改善心脏功能的能力。
与来自对照 MSC 的 EV 相比,此类 EV 可以转移更多的 miR-19a,从而在小鼠模型中恢复心脏收缩功能并减少梗塞面积。此外,缺氧诱导因子 1-α (HIF-1α) 转录因子通常在缺血期间稳定并上调多种心脏保护基因,这导致研究人员在牙科中突变 HIF-1α 基因(抗氧形式)用于治疗缺血相关疾病的牙髓来源的 MSCs。
与对照 EV 相比,来自过表达 HIF-1α 的 MSC 的 EV 具有增加的 EV 标记蛋白(如四跨膜蛋白)和增加的血管生成活性,这导致小鼠模型中心脏组织的修复增加。
一项类似的研究表明,源自过度表达 HIF-1α 的 BM-MSCs 的 EV 能够促进骨再生并减少类固醇诱导的股骨头缺血性坏死。诸如 Akt 之类的信号分子已在 MSCs 中被利用以提高其有效性。
来自过表达 Akt 的 UC-MSC 的 EV 比对照 EV 含有更高水平的 Akt,这会导致加速内皮细胞的增殖、迁移和血管形成,从而提高心脏修复的效率。这种作用是由促进缺血性心脏血管生成的内皮细胞中增强的 PDGF-D 产生介导的。在 CM 中观察到过表达 Akt 的 MSC-EV 的这些心脏保护作用的一些迹象,这些作用归因于分泌的卷曲相关蛋白。
间充质干细胞位于肿瘤细胞附近,据报道参与肿瘤进展。人们普遍认为,肿瘤微环境可以改变 MSC-EV 的含量,并导致它们趋向于更促肿瘤的表型。例如,科学家Roccaro 等人,表明来自多发性骨髓瘤患者的 BM-MSC-EVs 与来自正常健康受试者的 EVs 具有不同含量的肿瘤抑制 miRNAs,并且这些来自患者的 EVs 促进了多发性骨髓瘤肿瘤的生长,而来自健康个体的 EVs 抑制了肿瘤细胞的生长。
因此,还研究了 MSCs 的遗传修饰如何影响肿瘤生长。肿瘤坏死因子相关凋亡诱导配体 (TRAIL) 已被证明是一种有前途的癌症治疗药物,基于此,科学家Tuan 等人将该基因转染到 MSCs 中,然后测量源自这些细胞的 EVs 的癌细胞杀伤功效。
这种 EV 装饰有高表达的 TRAIL,并在各种癌细胞系中诱导细胞凋亡,但在原发性支气管细胞中没有。除了基因过表达外,肿瘤相关基因敲低的作用也在 BM-MSCs 中得到表征。来自 p53 缺陷的 BM-MSCs 的 EV 富含促进胃癌进展的 UBR2 蛋白。这种间接将 UBR2 靶向靶细胞的 p53 癌基因的调节通过调节 Wnt/β-连环蛋白途径增强了肿瘤的生长和转移。
b. MSCs中miRNA过表达
使用靶向转录和转录后调节的 miRNA 可能为治疗许多疾病提供一种新的选择。然而,miRNA 治疗的进展受到阻碍将 miRNA 递送至靶器官的障碍。EV 已成为传递 miRNA 的有效载体,因此许多研究人员一直在设计 MSC 以将 miRNA 加载到 EV 中并看到了有效的治疗效果(表 2)。
表2,在体外和体内模型中进行的基因转染MSC研究
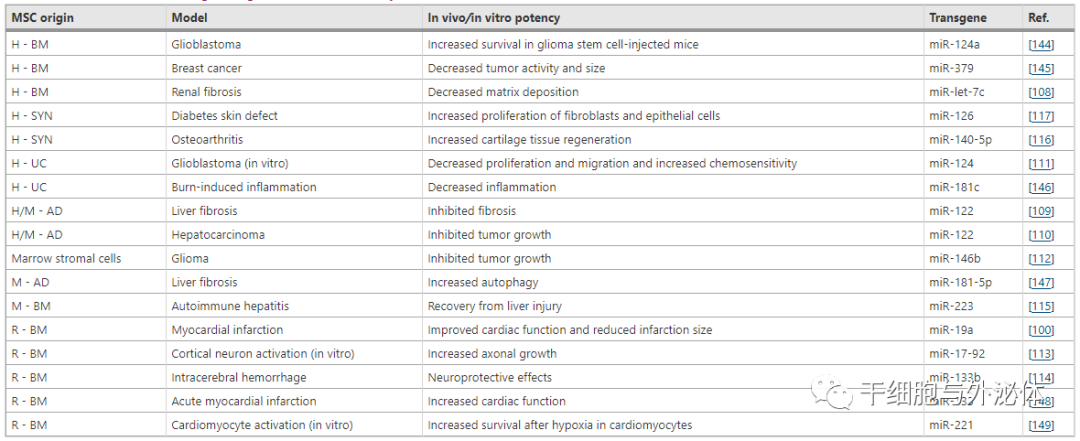
| 缩写:MSC 间充质干细胞、H 人、M 小鼠、R 大鼠、BM 骨髓、AD 脂肪组织、UC 脐带、SYN 滑膜
小鼠肾损伤和肝纤维化模型用于研究 EV 介导的 miR-let7c 和 miR-122 的抗纤维化作用。在这个小鼠模型中,从 MSCs 释放的 EVs 被设计为过度表达 miR-let7c,其中包含丰富的 miR-let7c,能够减轻肾损伤并显着下调 TGF-β1 和肾脏中下游纤维化基因的表达,因此提供了使用工程化 MSC 通过 EV 治疗性递送 miRNA 的主要例子。
鉴于 miR-122 通过负向调节肝细胞增殖在肝纤维化中起关键作用,miR-122 在 AD-MSCs 中被修饰以产生 miR-122 水平增加的 EV。这些 EV 通过 miR-122 诱导的靶基因(如胰岛素样生长因子受体 1、细胞周期蛋白 G-1 和脯氨酰-4-羟化酶 α-1)的下调介导 MSC 和肝星状细胞之间的通讯。
除了 EV 相关的 miR-122 的抗纤维化作用外,相同的含有 miR-122 的 EV 使肝细胞癌细胞对索拉非尼的化学治疗作用更敏感。与此一致,其他 miRNA 修饰已被证明赋予 MSC-EV 抗肿瘤作用。为了减轻将 miRNAs 靶向多形性胶质母细胞瘤的困难,脐带Wharton’s jelly源间充质干细胞(WJ-MSCs) 用 miR-124 过表达,衍生的 EVs 增强了对替莫唑胺的化学敏感性并减少了胶质母细胞瘤细胞的迁移。
在另一项研究中,过表达 miR-146b 的大鼠脑 MSC-EV 被用于减少胶质瘤异种移植物的肿瘤负荷,并且这些 EV 的肿瘤内给药减少了大鼠脑中胶质瘤的生长。
miR-17-92 和 miR-133 的神经保护活性已在来自表达 miRNA 的 MSCs 的 EV 中增强。从转染 miR-17-92 的 MSCs 中收获的 EV 显示出皮质神经元的轴突生长显着增加,其特征是与对照 EV 相比具有更高的轴突伸长速度。
在脑出血大鼠模型中,含有 miR-133 的 MSC-EV 能够产生有助于阻止神经元退化的促生存信号反应,这是通过抑制 RhoA 和激活 Erk172/cAMP 反应来介导的元件结合蛋白。
研究还检查了 MSC-EV 在不同疾病模型中传递的 miRNA 的再生作用。来自过表达 miR-223 的 BM-MSCs 的 EVs 用于自身免疫性肝炎的小鼠模型,EVs 可以通过 miR-223 诱导的靶细胞因子表达下调和含有 3 和 caspase-1 的 NLR pyrin 结构域的下调来预防肝损伤活动。
此外,一项对源自过表达 miR-140-5p 的人类滑膜 MSCs 的 EV 的研究表明,在大鼠模型中增强了软骨组织再生并减少了膝关节骨关节炎,并且源自过表达 miR-126 的人类滑膜 MSCs 的 EVs 愈合了糖尿病大鼠模型中的全层皮肤缺损。
结论和观点
关于EV 成分与 EV 生物效应之间的关系,最常见的分子是蛋白质和 miRNA。已经研究了各种用特定蛋白质和核酸外源加载分离的 EV 的策略,例如,电穿孔、冻融循环、皂苷介导的加载和低渗透析。
此外,许多团体已经开始使用转基因 MSCs 将所需的货物包装在 EV 中,这些转基因 MSCs 经过基因改造以过度表达某些蛋白质和 miRNA。然而,这需要优化条件,以使转基因细胞外囊泡获得更有效的功能特性。此外,还需要对 EV 成分的质量控制以及这些 EV 的安全性和有效性进行全面研究,然后才能将其用于临床应用。
在讨论 MSC-EV 相对于 MSC 的优势之前,必须考虑对以下问题进行仔细调查的必要性。许多临床试验已经描述了各种外部因素对 MSCs 特性的影响。
供体变异、衰老、细胞培养方法和免疫原性的细微差异被证明会导致 MSC 治疗的功能改变。例如,经历细胞衰老的 MSCs 会促进代谢功能障碍并失去其间充质可塑性和抗炎作用,这可能导致 MSC 治疗失败。据作者所知,没有研究评估 EV 治疗活性与 MSC 衰老之间的关系。然而,在对 EVs 进行功能评估时,还需要进一步研究衰老细胞的含量,深入了解相关机制将有助于成功开发 MSC-EVs 用于临床。
总之,MSCs 通过各种囊泡成分以及细胞本身及其分泌的可溶性因子具有潜在的治疗功能,并且 MSCs 可以进行修饰以提高其产生的 EV 的数量和有效性。因此,MSC 衍生的 EV 可以用作强大的治疗剂,在许多不同的疾病中提供抗炎和再生化合物。未来的工作将集中在开发生物工程 MSC,以显着提高 EV 的产量,从而安全地转移各种有效的治疗分子。
编辑:小果果,转载请注明出处:https://www.cells88.com/cells/gxb/9288.html
免责声明:本站所转载文章来源于其他平台,主要目的在于分享行业相关知识,传递当前最新资讯。图片、文章版权均属于原作者所有,如有侵权,请及时告知,我们会在24小时内删除相关信息。
说明:本站所发布的案例均摘录于文献,仅用于科普干细胞与再生医学相关知识,不作为医疗建议。
 微信扫一扫
微信扫一扫  支付宝扫一扫
支付宝扫一扫 