

一、微血管系统 — 二、基底膜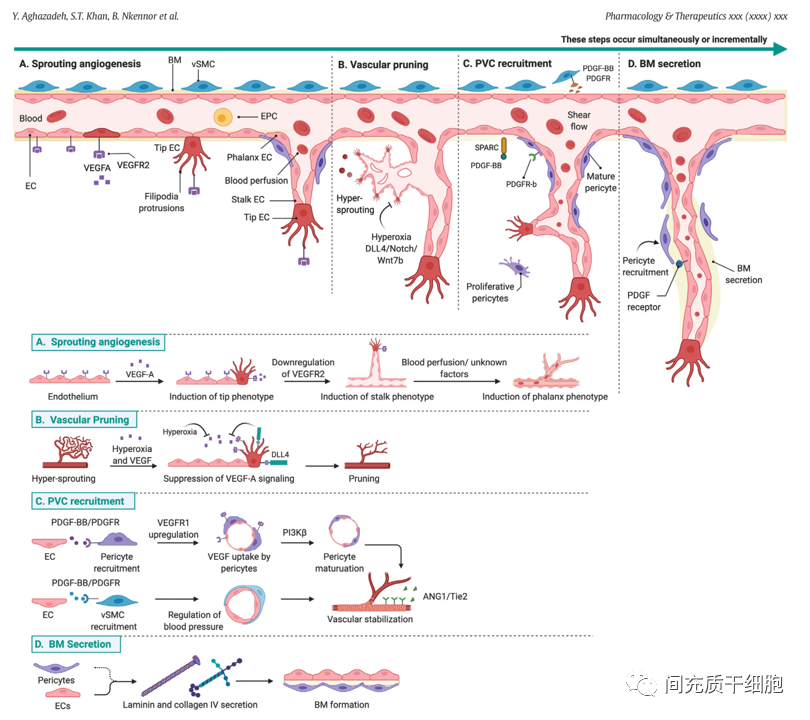

2.5被认为是冠状动脉微血管功能障碍(CMD)[17]。皮肤微血管功能可以通过各种非侵袭性到轻度侵袭性的临床评估来衡量[18]。
基底膜是血管系统的脱细胞成分,富含多种功能蛋白,由内皮细胞和血管周细胞分泌,对血管的完整性和稳定性至关重要。基底膜支持内皮细胞和血管周细胞之间的相互作用,并为血管和周围非血管组织提供机械信号。基底膜已被鉴定出40多种基底膜蛋白质,它们的组合/比率在不同组织之间存在显著差异[20]。
三、血管系统的关键细胞类型
(1)内皮细胞
血管周围细胞的功能正常有助于血管完整性和屏障功能[24]。内皮细胞是血管周围细胞群体中最重要的一种细胞类型。内皮细胞的α-平滑肌肌动蛋白(α-SMA)参与了血管收缩和血流调节。在静止状态下,内皮细胞也需要线粒体产生ATP的能量来维持生理功能和稳态。内皮细胞中大多数ATP是通过糖酵解代谢产生的,虽然内皮细胞的位置靠近血流并与足够的氧气接触[25]。在内皮细胞糖酵解产生ATP的生理过程中,内皮细胞的生长和增殖伴随着糖酵解代谢的增加,糖酵解代谢的增加又依赖于PFKFB3活性的上调,所以PFKFB3酶活性对调节内皮细胞的糖酵解至关重要[26-28]。
原代内皮细胞具有较高的免疫原性,异体移植治疗能引起免疫排斥,因而需要搭配免疫抑制药品的使用[29]。不过,大血管内皮细胞不表达MHC-II类分子,因此其免疫原性低于微血管内皮细胞[30]。再生研究中使用的血管细胞最常见的来源是来自人体组织的原代内皮细胞。原代人内皮细胞可从活组织或其他可获得的组织中获得,如脐带静脉和脂肪组织,并在体外扩增几代。人脐静脉内皮细胞(HUVEC)于1974年首次分离和培养[31],是临床前研究中最常用的内皮细胞来源。因为内皮细胞根据血管结构树的空间定位显示出独特的生理和转录组学特征[32, 33],因而人脐静脉内皮细胞是从大血管静脉获得的,它们可能不适合生成微血管或动脉,因此,人脐静脉内皮细胞和其他原代内皮细胞在临床上的应用面临着一些困难。
(2)周细胞(间充质干细胞)
周细胞嵌入血管基底层,包裹小直径微血管,如毛细血管前小动脉、毛细血管和毛细血管后小静脉。周细胞参与新生血管是通过内皮细胞和细胞外基质(ECM)的多种信号控制的,包括PDGF-BB和SPARC[39]。成熟周细胞通过表达VEGFR1来接收内皮细胞分泌的VEGF信号来调节血管生成,从而诱导内皮细胞出现茎表型并导致出芽方式的血管延长[40]。对成年小鼠的研究表明,周细胞数量的减少会导致出血和毛细血管过度扩张[41],并与糖尿病视网膜病变[42]和癌症[43]等疾病相关。
(3)血管平滑肌细胞
在血管结构层次(动脉和静脉、小动脉和小静脉)中,血管平滑肌细胞(vSMC)是主要的血管周的壁细胞类型。血管平滑肌细胞呈梭形,通过细胞内丰富的αSMA来控制细胞的收缩功能,从而调节局部血压[50, 51]。内皮细胞通过PDGF-BB信号与血管平滑肌细胞上的受体结合来吸引和黏附(招募)血管平滑肌细胞和周细胞[52, 53]。周细胞和血管平滑肌细胞分泌ANG1,激活内皮细胞表面的Tie-2受体,通过加强内皮细胞细胞间的相互作用来稳定血管;如果Tie受体出现突变,那么可以导致斑块状血管平滑肌细胞覆盖和血管畸形,从而破坏血管的稳定性[54, 55]。
(4)其他细胞类型
除了周细胞和血管平滑肌细胞外,还有一些其他类型的细胞对内皮细胞起支持作用,如成纤维细胞[56]、中性粒细胞[57]和巨噬细胞[58]。在小鼠下肢缺血的实验模型中,缺血周边局部组织有多个明显不同的成纤维细胞亚群(转录谱不同),而这些成纤维细胞亚群的存在有助于血管生成反应,而且还需要依赖于天然免疫信号的激活,才能有助于恢复缺血组织的灌注和恢复[59]。
四、血管病变的MSC治疗
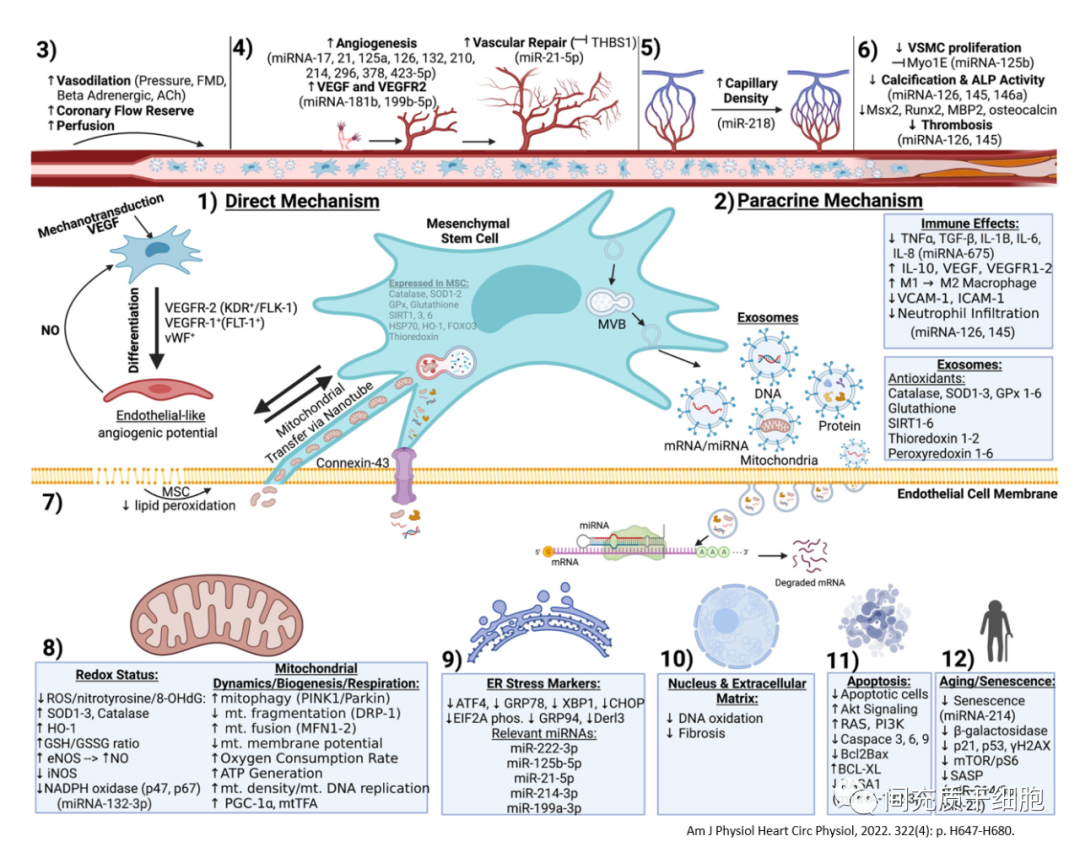
MSC通过分泌一些有功效的物质对受损组织发挥修复作用,或激活受损组织本身的干/祖细胞的激活和分化,即旁分泌作用机制,而不是MSC植入受损组织进行分化为组织细胞[60]。
—
(1)MSC的外泌体机制
MSC外泌体也可用于修复血管屏障功能,从而有利于治疗血管屏障障碍相关的疾病[61]。在MSC外泌体的内含物中,miRNAs是非常重要的具有细胞生物学活性的作用物质之一。MiRNAs可作为基因表达的抑制子,与特定mRNA的3′-非翻译(3′-UTR)区域结合,可通过促进mRNA降解或抑制mRNA翻译为蛋白质,在共同途径内同时调节多个基因[62, 63]。

—
MSC分泌的外泌体中,同时存在促进血管生成的miRNAs和抑制血管生成的miRNAs,不过抑制血管生成的miRNAs含量很低[64]。比如,miR-93通过作用于多个调节基因,增强内皮细胞增殖和减少骨骼肌细胞凋亡,从而促进后肢缺血后的毛细血管密度和灌注恢复[65]。外泌体所含的miR-125a可通过靶向抑制delta-like 4(DLL4)的表达,从而解除了delta-like 4对血管生成的抑制作用,进而促进内皮细胞的血管生成[66]。又如,miR-15a[67]和miR-503[68]同样为内皮细胞增殖和血管生成的负性调节因子。miR-92a和miR-100靶向对应于几种促血管生成蛋白的mRNAs,在肢体缺血和心肌梗死的小鼠模型中,抑制miR-19[69]、miR-92a[70]和miR-100[71]的活性可促进血管生长和受损组织的功能恢复。抑制可改善老龄小鼠缺血后的血流恢复,并减少对这些靶点的抑制。
(2)MSC的抗氧化作用
MSC介导的血管再生的一个主要因素是宿主组织的抗氧化作用[77, 78]。MSC对氧化应激具有高度抵抗力,因为它们组成性地表达过氧化氢酶、超氧化物歧化酶1-3(SOD1-3)、谷胱甘肽过氧化物酶(GPx)、SIRT 1/3/6,以及硫氧还蛋白、血红素加氧酶-1,谷胱甘肽(GSH)。这些抗氧化物质通过降低氧化应激、减少炎症以及增加宿主内皮细胞/血管平滑肌细胞抗氧化蛋白含量和/或活性,介导血管损伤和疾病的恢复[79-81]。
(3)MSC的细胞因子的作用
在MSC的培养上清中,MSC分泌多种可溶性因子-包括VEGF、VEGFR2、Ang-1/Tie2、单核细胞趋化蛋白-1(MCP-1)、巨噬细胞炎性蛋白-1α(MIP-1α)、MIP-1β、单因子、碱性成纤维细胞生长因子(bFGF)、PDGF、IL-1β、IL-10、干细胞衍生因子(SDF)-1、肝细胞生长因子(HGF)、胰岛素样生长因子1(IGF-1)、胸腺肽-β4、Wnt5a 等[60, 84]。
—
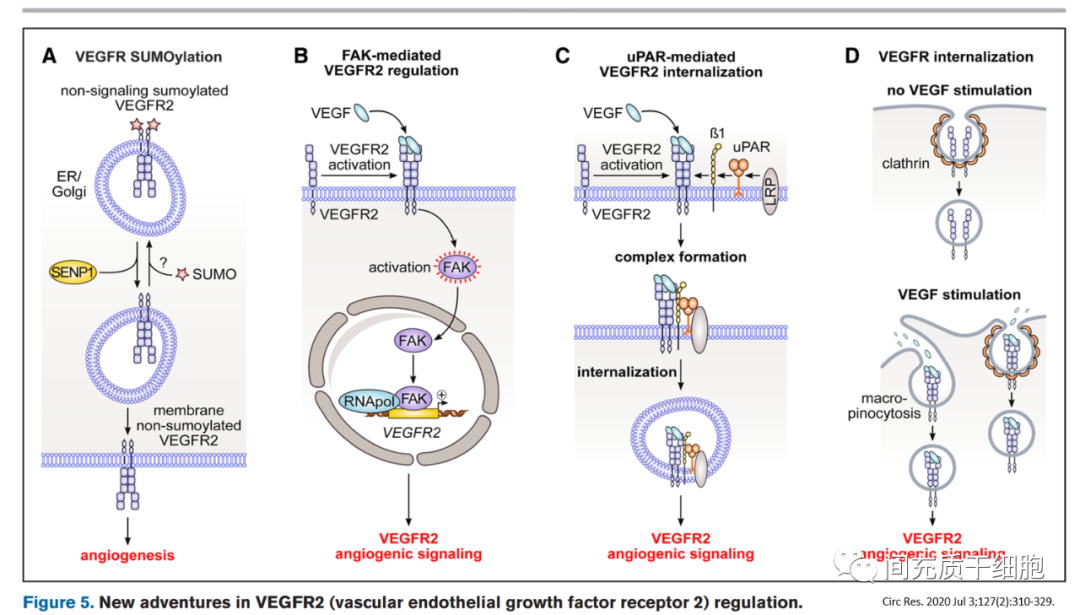
在缺氧和血清饥饿状态下,向内皮细胞中添加VEGF165a可通过减少细胞死亡和增加内皮细胞增殖以及增加糖酵解来诱导血管生成,但存在血管通透性和血管渗漏的现象[89]。而且人体临床研究使用VEGF165a等药物促进缺血区域血管生长以重建血供的治疗效果不佳[90]。
(4)其他机制
MSC来源的外泌体能改善受损细胞的线粒体功能,减少线粒体DNA的损伤和提高线粒体NDA的稳定性[91-93]。甚至在内皮细胞发生缺血/再灌注损伤的实验中,可以观察到MSC的线粒体可以直接从隧道纳米管转移给内皮细胞,增加内皮细胞产生更多的能量ATP,从而挽救损伤的内皮细胞[94, 95]。在脂多糖诱导的急性肺损伤模型中,肺泡上皮细胞活性氧增加、线粒体膜电位降低和ATP产生降低,肺部血管屏障功能减弱;MSC与肺泡上皮形成了含有连接蛋白43(Cx43)的缝隙连接通道,释放出被上皮细胞吞噬的含有线粒体的外囊泡(外泌体),线粒体从MSC转移到肺泡上皮细胞导致肺泡ATP浓度增加,减少了LPS诱导的急性肺损伤炎症[96]。
五、小结
MSC治疗也可以在高血压和外周阻力增加的情况下促进微血管再生。比如在慢性肾动脉狭窄大鼠模型中,MSC治疗促进血管紧张素2型受体表达增加、恢复狭窄肾脏的微血管树状网络结构,从而提高肾小球滤过率和肾血浆流量[100]。MSC能恢复肝脏微血管功能和降低门静脉高压力[101]。然而,自体干细胞疗法的持续给药受到患者可获取数量的限制,以及捐赠或接受细胞疗法的患者是否存在可能影响细胞疗法的病理状况[102]。
it is naive to think that one administration of cells is enough to provide
life-long therapeutic remedy.)

—
简而言之,在基于细胞的再生医学中,想在体内重建成熟的具有功能的网状结构血管,仍然是有待突破的科学难题。单细胞表观基因组学和转录组学的检测技术有可能分析细胞随时间变化的轨迹,这无疑将有助于确定血管再生过程中血管成熟的关键细胞生物学变化。相信随着对血管再生各个环节的深刻理解,借助于MSC和多种细胞的相互作用,将来是可以完美地解决血管生成的难题。
-END-
来源:间充质干细胞
编辑:小果果,转载请注明出处:https://www.cells88.com/cells/gxb/33668.html
免责声明:本站所转载文章来源于其他平台,主要目的在于分享行业相关知识,传递当前最新资讯。图片、文章版权均属于原作者所有,如有侵权,请及时告知,我们会在24小时内删除相关信息。
说明:本站所发布的案例均摘录于文献,仅用于科普干细胞与再生医学相关知识,不作为医疗建议。
 微信扫一扫
微信扫一扫  支付宝扫一扫
支付宝扫一扫 