


前言 /
间充质干细胞(MSC)广泛存在于人体中,而且伴随着人体的整个生命周期。肿瘤发病率越来越高,而且肿瘤细胞的生长速度远远超过机体的正常细胞。那么,面临的一个问题就是, MSC和肿瘤细胞有什么样的相互作用?本文就这个话题深入讨论几个相关知识点。
广泛存在于人体的间充质干细胞
1990年Diaz-Flores博士发现骨骼和骨骼周围组织血管周围有一种细胞具有祖细胞的特性,能促进血管新生和组织修复的功能,鉴定为血管周细胞(pericytes)[1];随后发现这种周细胞具有向成骨细胞和软骨细胞分化的能力[2, 3]。脂肪组织血管的周细胞同样和MSC具有相似的基因谱[4]。从各种组织中分离出来的周细胞产生的MSC,都具有相同的细胞表面抗原及其体外多能性[5]。小鼠实验证明MSC分布在全身血管周围,因此几乎全身所有的器官和组织基质中均存在MSC[6]。重要的是,MSC可以从每个血管化的组织中分离出来,甚至可以从月经中分离出来(即破碎的血管释放血管周细胞)[7-9]。因此Caplan教授提出大多数间充质干细胞是从血管周围或壁细胞(周细胞)分化而来的[10]。
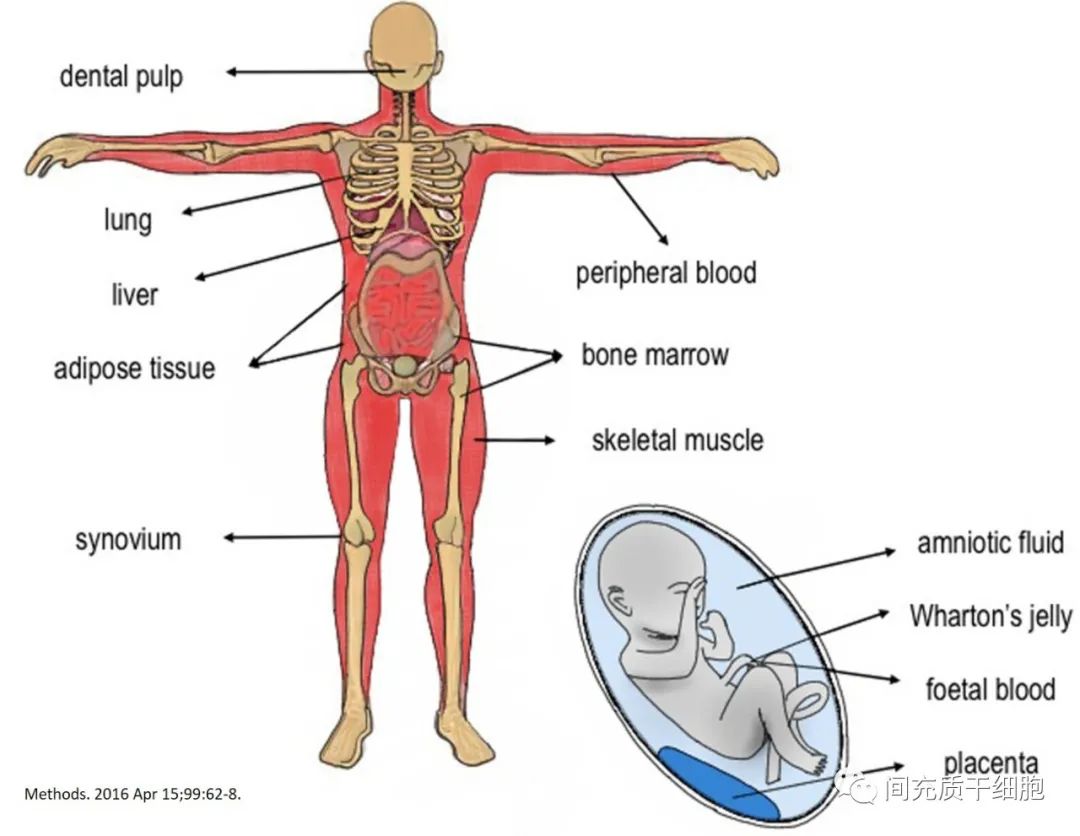
所以,基本上有大血管存在的地方,都会有MSC的身影。当然了,骨髓中存在含量更丰富的MSC。
肿瘤组织中的间充质干细胞
肿瘤细胞需要不断增殖,那就需要有大量的血液供应,所以肿瘤组织内部出现血管的生成,自然伴随着大量的周细胞(或MSC)来参与组建血管。1999年,Maestroni等在肺癌和黑色素瘤的肿瘤部位找到MSC[11]。随后,MSC被证明存在于多种肿瘤组织基质中,包括黑色素瘤[12]、胰腺癌[13]、卵巢癌[14]、结肠癌[15]、乳腺癌[14]、肺癌[16]、恶性胶质瘤[17]和Kaposi肉瘤[18]。
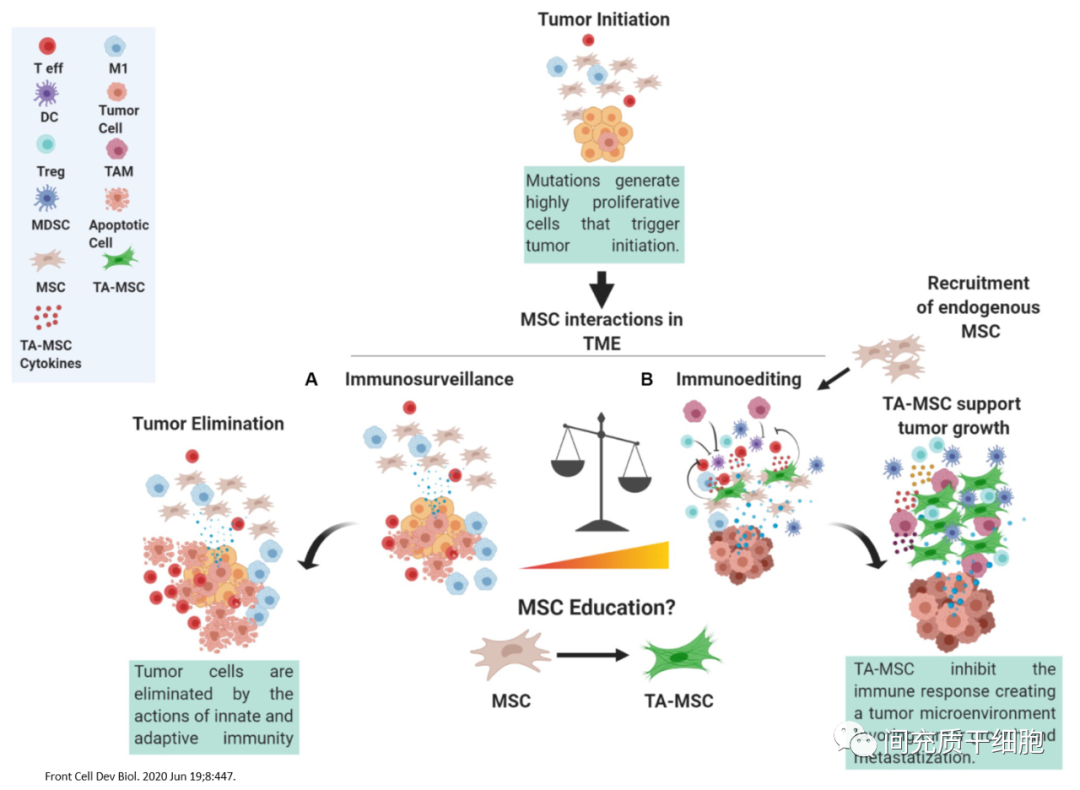
另外,MSC具有趋化到炎症部位的本能,而肿瘤具有“不能治愈的损伤”(慢性炎症)的特性[19],此特性能吸引体内的MSC趋化到肿瘤部位[20, 21]。包括机体里在骨髓和脂肪中的MSC,均会被募集趋化到肿瘤的部位[22, 23]。
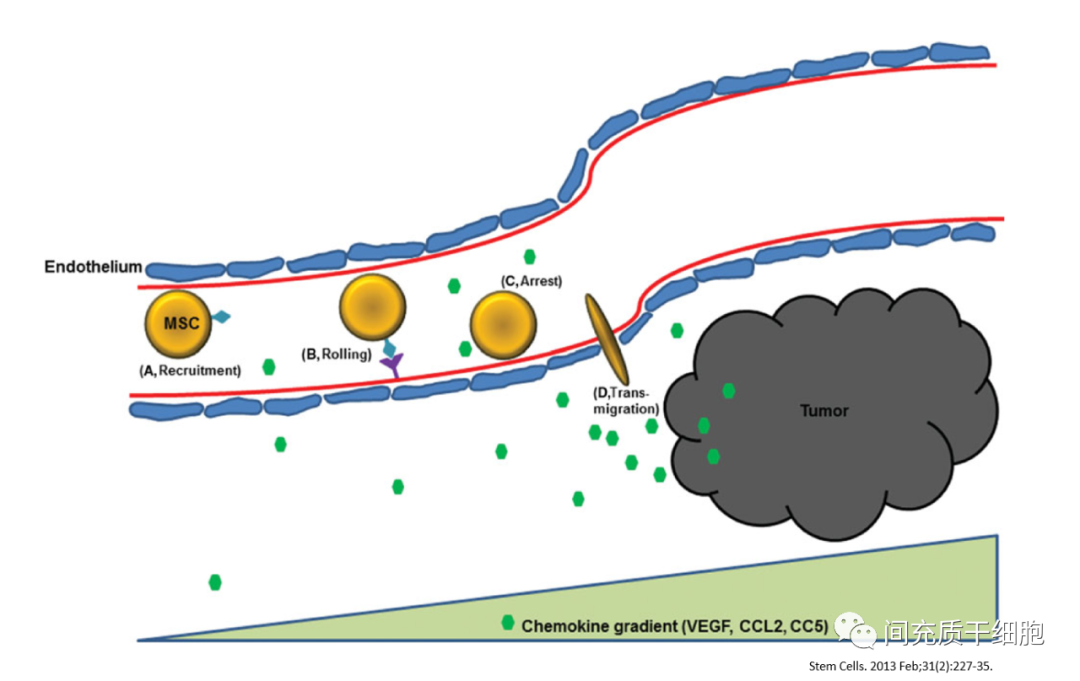
因此,在肿瘤组织里发现MSC的存在,更可能是MSC在尝试消除肿瘤部位的慢性炎症,只是限于数量和某种功能的缺失,导致不能有效地清除这个慢性炎症,反而会促进了肿瘤细胞的增殖。
间充质干细胞不会转变为肿瘤细胞
目前认为肿瘤干细胞并不是来源于正常的干细胞,而可能是肿瘤细胞在生存压力下的进化突变结果[24-26]。肿瘤干细胞在自己的巢(niche)里维持自身的更新和持续分化为肿瘤细胞。不仅正常健康的MSC不具有形成肿瘤的能力[27, 28],即使是慢性粒细胞性白血病患者骨髓的MSC,也不具有形成肿瘤的能力[29]。其实很好理解这点,如果正常健康的MSC能形成肿瘤,那么每一个人从出生开始,就伴随着肿瘤的生长,然而事实上并非如此。
既然人体内存在的MSC不具有成瘤性,那么MSC经过体外扩增培养后,是否具有成瘤性?研究发现,人MSC经过体外培养低代数扩增后,也不具有成瘤性,但高代数扩增能引起MSC染色体异常[30-32]。骨髓MSC在培养至18代的时候出现了染色体异常和端粒酶缩短[31],而脐带MSC在培养至30代才会出现染色体异常[32]。但是一般都会应用10代前的MSC,所以不用担心染色体异常的问题。
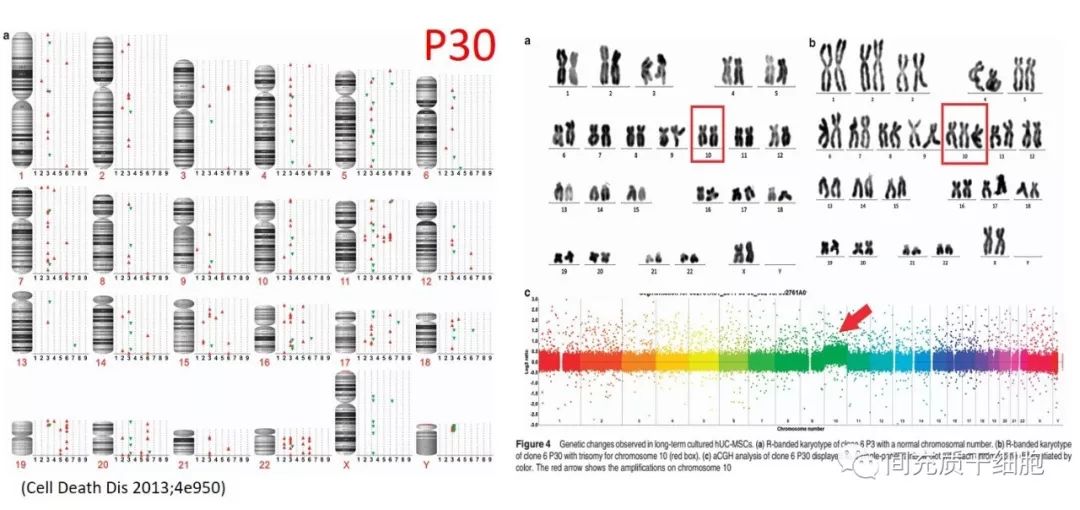
2010年的一篇由法国13个科研单位共同发表在《血液》杂志的实验文章,发现人骨髓MSC在P1代的时候就可以出现染色体异常(非整倍数),但是染色体异常并不影响到MSC的倍增时间;而且小鼠成瘤实验证明,即使MSC的染色体异常和长期培养导致的衰老,移植到小鼠体内后并不能成瘤[33]。
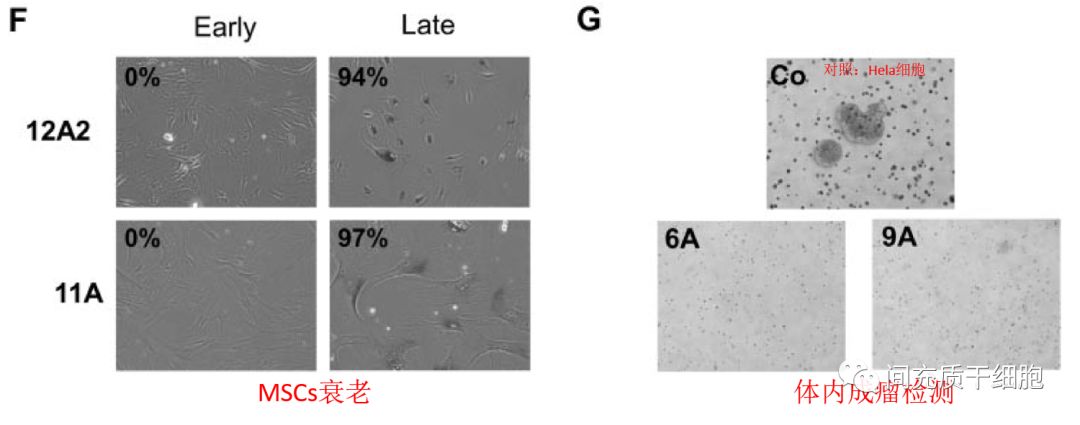
实验证明染色体异常的MSC也不具有成瘤性,但是体外培养导致的染色体异常和MSC衰老密切相关。衰老的人骨髓MSC的功能变化主要与细胞发育、细胞生长/增殖、细胞死亡、细胞信号转导/相互作用和细胞运动有关[34]。所以,临床应用的时候,还是选择年轻态的MSC更合适。
间充质干细胞促进 或抑制肿瘤的矛盾结果
MSC和肿瘤的相互作用充满了争议。一些研究显示出MSC的抑瘤作用,而另一些研究则显示了MSC对肿瘤的促进增殖作用。
MSC可能通过分化为周细胞或内皮细胞直接支持肿瘤血管形成,也可能间接通过辅助促血管生成因子的分泌来支持肿瘤血管形成[35, 36]。在小鼠和人类器官的血管周围组织中已经发现了大量的MSC样细胞[5, 37]。周细胞在血管稳定中起着重要作用,但MSC也可以分化为内皮细胞,这可能会增加血管密度和新生血管[38]。因此,一些动物实验证明MSC促进肺癌[39, 40]、乳腺癌细胞[41]、卵巢癌细胞[42]、黑色素瘤细胞[43]和胃癌细胞[44]的生长。观察到MSC促进肿瘤的生长,推测其原因可能在于MSC分泌生子因子促进肿瘤细胞的生长、促进肿瘤组织血管的生成、参与了肿瘤干细胞生子微环境的构建(niche)[45, 46]。
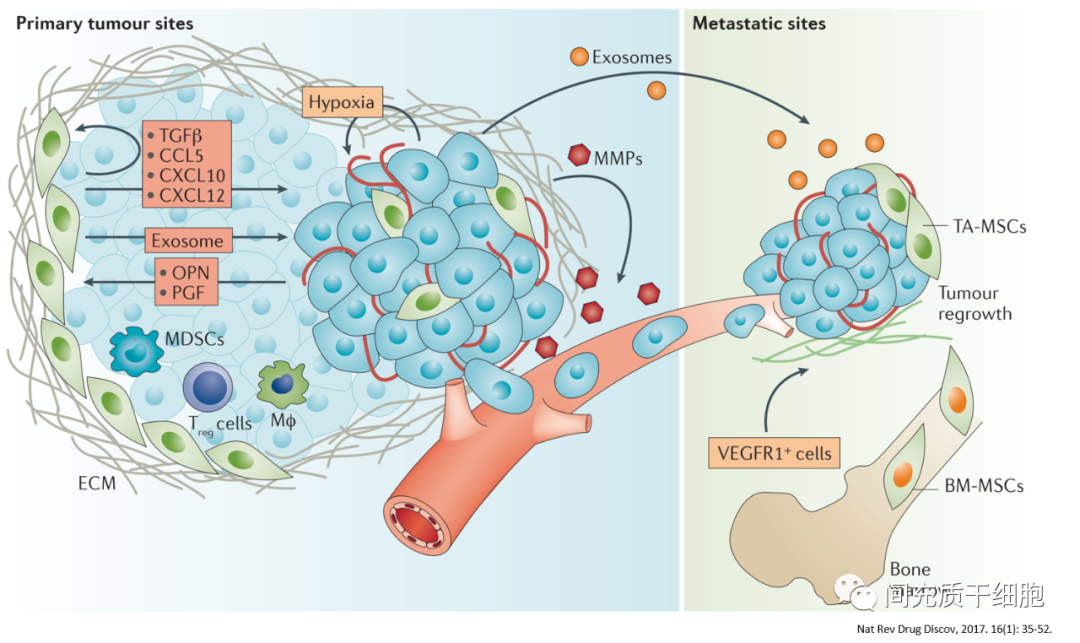
另外,也有一些动物模型上也显示出了抗肿瘤活性,比如Kaposi’s肉瘤[18]、Lewis肺癌和恶性黑色素瘤[11]、结肠癌[47]。MSC与Kaposi’s肉瘤细胞相互接触,通过抑制Akt活性,下调Akt的表达,从而抑制了Kaposi’s肉瘤的生长[18]。联合使用TNF-α、或TGF-β、或IFN-α/β、或内毒素,都没有影响到MSC对Lewis肺癌和恶性黑色素瘤的抗肿瘤活性[11]。MSC的培养上清液(富含生长因子)能下调肝癌细胞和乳腺癌细胞的NFκB的表达水平,从而抑制肝癌细胞和乳腺癌细胞的生长[48]。MSC抗肿瘤的活性可能与下调肿瘤细胞的Akt、NFκB和Wnt信号通路相关[49, 50]。
在血液系统恶性肿瘤中,同样存在相互矛盾的研究,MSC分别通过抑制肿瘤细胞的增殖或凋亡来抑制或促进肿瘤生长[51]。比如,MSC促进了急性淋巴细胞白血病株Reh细胞的增殖[52],但是又抑制了淋巴瘤细胞株的增殖[53, 54]。
MSC在治疗肿瘤方面的作用包括抑制血管生长[55, 56]、通过阻止细胞周期来降低细胞增殖[50, 57, 58]。然而,MSC也被认为支持肿瘤血管形成[36, 59],发挥免疫抑制的作用[60, 61],增加肿瘤转移和复发率[41, 62-65],甚至导致化疗耐药[66-68]。从目前相互矛盾的研究结果来看,MSC似乎可以影响既能抑制增殖又能抑制凋亡的通路,MSC的双重角色可谓一把“双刃剑”[69, 70]。

基于MSC在肿瘤领域的研究结果表现出相反的差异,这可能是由于非标准化的实验方法,缺乏特异性的细胞表面标记来识别MSC亚群,以及MSC的异质性很容易受到周围环境的影响[51]。有学者提出,MSC的来源可能决定了其对肿瘤细胞的反应[49]。脂肪来源的MSC通过分泌VEGF促进黑色素瘤细胞的生长,然而对胶质母细胞瘤细胞却没有促生长的作用[71]。但是,也有研究发现脂肪来源的MSC促进胶质母细胞瘤的生长,而脐带来源的MSC通过提高TRAIL的分泌而抑制其生长[72, 73]。另外,实验研究所用的肿瘤细胞系也可能会影响了MSC对肿瘤细胞抑制或促进的作用效果[71]。
在一项血液恶性肿瘤的先导性临床研究中,随机分为两组,其中一组患者接受来自人类白细胞抗原相合的同胞供者的造血干细胞(HSC),而另一组患者接受HSC与MSC联合移植;虽然MSC可预防移植物抗宿主病(GVHD),但恶性血液病的复发率高于对照组[74]。
间充质干细胞作为药物运送工具
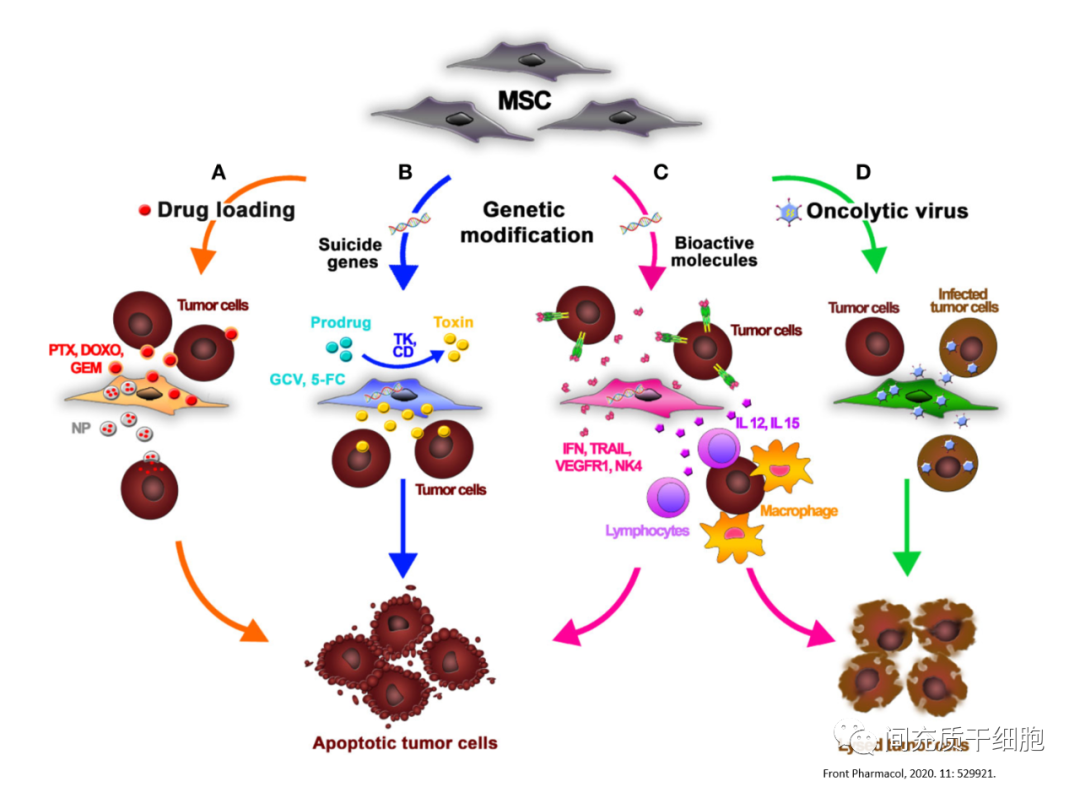
由于MSC具有低免疫原性和趋化到肿瘤基质的特性,是一个非常理想的携带抗肿瘤生物活性药物的载体[12]。2002年首次被用来传递干扰素-β,用于治疗黑色素瘤[12],减少肿瘤生长,延长小鼠模型的生存期。从那时起,研究人员相续基因改造MSC用于各种肿瘤的治疗[75],比如转染IFN-β治疗胰腺癌[13]和黑色素瘤[76]、转染IL-12治疗黑色素瘤和肝癌[77]及胶质瘤[78]、转染IFN-α治疗黑色素瘤[79]、转染IFN-γ治疗白血病[80]、IL-12治疗黑色素瘤、肺癌和肝癌[81]、IL-24治疗肺癌[82]、转染NK4治疗肺癌[83]、转染TRAIL治疗乳腺癌[16]和胶质瘤[17]及肺癌[84, 85]、转染溶解瘤细胞的腺病毒治疗卵巢癌[86]。
MSC可以携带传统的抗癌细胞毒药,均匀地浸润肿瘤组织,并改善治疗剂有效载荷在肿瘤内的分布[87, 88]。比如,携带抗癌药物紫杉醇(PTX)的纳米工程MSC可以将细胞中的紫杉醇转移到肿瘤部位,并在几天内释放药物,肿瘤细胞增殖减少,抑制血管生成,并增加肿瘤基质内的凋亡,提高了肿瘤小鼠的存活率,尽管MSC携带的紫杉醇的剂量明显较低[89, 90]。MSC与紫杉醇还可以通过挤压混合后,采用挤压法和超速离心法分离MSC的胞外囊泡模拟物(MSC-EM),这些载有紫杉醇的MSC-EM在体外和体内都显示出对乳腺癌的治疗效果[91]。MSC分泌的胞外囊泡包裹阿霉素,单次静脉注射即可显著抑制结肠癌生长[92]。
间充质干细胞在临床肿瘤治疗中的 辅助作用
放化疗的一个常见的副作用就是口腔和胃肠道的溃疡,影响了营养的吸收,同时降低患者的生活质量[93, 94]。给大鼠腹腔放射性照射(放疗)后,大鼠出现腹泻、血便、肠粘连、肠溃疡和疤痕组织增生、肠黏膜的绒毛组织受到严重破坏而不能自我恢复;但是,给予MSC治疗后,肠绒毛组织修复良好、溃疡愈合,而且不出现肠粘连,大鼠的存活率明显提高[95]。注射的MSC也能迁移到照射部位,促进放射性照射导致的皮肤烧灼伤的愈合[96]。
骨髓抑制也是化疗的常见毒副作用之一,而且威胁到生命,关系到化疗的成功与否,急需对症处理[97, 98]。1995年,从恶性血液病患者骨髓抽取并分离培养出MSC,然后回输到患者体内,临床观察到促进肿瘤患者的造血干细胞的造血机能,改善贫血,而且提高肿瘤患者的生存质量(改善睡眠和饮食,减轻疼痛)[99]。由于MSC分泌多种营养因子及G-CSF、M-CSF、GM-CSF,体外实验证明MSC能促进造血干细胞的造血克隆形成能力[100, 101],这提示MSC能提高患者的骨髓造血能力[102, 103]。动物实验更是直接证明MSC能够恢复受损后的骨髓基质功能,促进骨髓损伤后的造血恢复[103]。
化疗药物(环磷酰胺)损伤大鼠卵巢,导致大鼠卵巢功能受损(卵巢滤泡变小),给予腹腔注射MSC后,通过旁分泌机制,分泌的细胞生长因子(VEGF、IGF-1、HGF)直接修复损伤的卵巢组织,使得损伤的卵巢功能得到恢复[104-106]。MSC也能阻断化疗药物(白消安)对雄性大鼠精子发生功能的损伤,有效地恢复雄性大鼠的生育功能[107, 108]。Pinarli等建立化疗药物(阿霉素)导致的心脏毒性大鼠模型,然后给予MSC治疗,发现MSC既能预防化疗药物导致的心脏毒性反应,又能治疗已经出现的心脏毒性反应[109]。
接受化疗的患者,肿瘤细胞反弹性生长,常常导致化疗的失败和病情的恶化。但是化疗药物在杀伤肿瘤细胞的同时,也同样杀伤了机体里的MSC。紫杉醇、长春新碱、依托泊苷和阿糖胞苷能损伤MSC,诱导其凋亡;只是长春新碱和地塞米松对MSC的轻度损伤是可以恢复的[110]。急性淋巴细胞白血病儿童患者接受一定剂量的化疗,损伤骨髓MSC的支持造血功能[111]。接受常规剂量的化疗并不影响MSC的增殖能力[112],但是接受高剂量化疗的患者,其骨髓来源的MSC的克隆形成能力和增殖能力均降低[113]。因此,需要深入研究和评价癌症患者接受多次化疗后,除了免疫细胞的损伤,肿瘤细胞反弹性生长与MSC的损伤有没有关系?
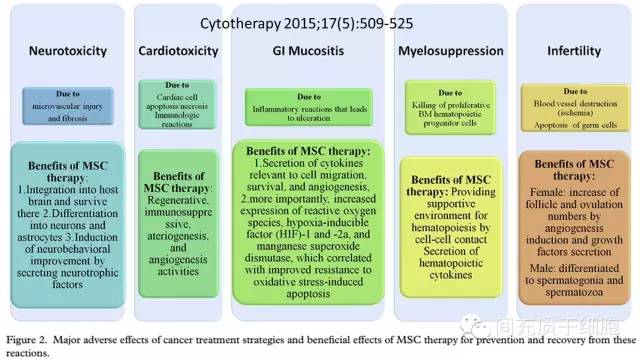
由于健康的MSC具有促进组织器官修复再生的功能、抗炎的特性、分泌众多营养性细胞因子等特点,使得健康的MSC成为理想的肿瘤/癌症治疗的辅助手段,有利于治疗抵消放化疗过程中伴随的毒副作用,既能促进机体的恢复,又能提高放化疗的效果[114]。
小结
在讨论MSC与肿瘤的关系的时候,需要非常注意和谨慎对待动物肿瘤模型给我们带来的知识,因为动物肿瘤模型的建立,需要用到免疫缺陷动物,而没有免疫系统的存在,MSC分泌大量的营养性细胞因子,难免会促进某些肿瘤细胞的生长。在一个靶向性很好的抗肿瘤药物,由于其不会出现和免疫细胞的相互作用,这种情况下,可以采用免疫缺陷动物建立的肿瘤模型来评价治疗效果。免疫系统的存在与否,严重影响到MSC对肿瘤的作用效果。但是,在人体内,由于免疫系统的存在,需要评估MSC通过调节免疫细胞来影响到其对肿瘤细胞的作用。
基于MSC的免疫调节作用,有专家认为MSC可以作为肿瘤组织的干预靶点[115, 116]。但是如果选择MSC作为治疗肿瘤的干预靶点,需要非常精准的区分肿瘤组织的MSC和其他部位健康的MSC,以免误伤友军。
由于MSC具有很强的支持造血、消炎、分泌多种生长因子而促进组织器官的自我修复等作用,理论上能缓解和对抗肿瘤的放化疗引起的毒副作用。动物实验显示MSC能通过多种作用机制修复放疗导致的健康组织的损伤,抵消放疗的毒副作用,延长放疗后动物的生存时间。但是,目前没有临床(人体)研究数据证明MSC促进肿瘤的生长还是抑制肿瘤的生长,也未见有MSC联合放化疗治疗肿瘤的临床结果报道,这需要今后临床严谨的科学研究。
将健康有活性的间充质干细胞作为一个治疗手段,应用于肿瘤患者,也许还有争议;但是在肿瘤患者的临终关怀阶段,健康有活性的间充质干细胞能提高生存质量,减轻痛苦,能让肿瘤患者有尊严地走完人生屈指可数的最后一段时光。
相信MSC在将来的临床研究中将大放异彩!
-END-
来源:间充质干细胞
编辑:小果果,转载请注明出处:https://www.cells88.com/cells/gxb/33143.html
免责声明:本站所转载文章来源于其他平台,主要目的在于分享行业相关知识,传递当前最新资讯。图片、文章版权均属于原作者所有,如有侵权,请及时告知,我们会在24小时内删除相关信息。
说明:本站所发布的案例均摘录于文献,仅用于科普干细胞与再生医学相关知识,不作为医疗建议。
 微信扫一扫
微信扫一扫  支付宝扫一扫
支付宝扫一扫 